The central nervous system (CNS) and the immune system collectively coordinate cellular functionalities, sharing common developmental mechanisms. Immunity-related molecules exert an influence on brain development, challenging the conventional view of the brain as immune-privileged. Chronic inflammation emerges as a key player in the pathophysiology of Alzheimer's disease (AD), with increased stress contributing to the disease progression and potentially exacerbating existing symptoms. In this study, the most significant gene signatures from selected RNA-sequencing (RNA-seq) data from AD patients and healthy individuals were obtained and a functional analysis and biological interpretation was conducted, including network and pathway enrichment analysis. Important evidence was reported, such as enrichment in immune system responses and antigen processes, as well as positive regulation of T-cell mediated cytotoxicity and endogenous and exogenous peptide antigen, thus indicating neuroinflammation and immune response participation in disease progression. These findings suggest a disturbance in the immune infiltration of the peripheral immune environment, providing new challenges to explore key biological processes from a molecular perspective that strongly participate in AD development.
Citation: Dimitra Anatolou, Marios G. Krokidis. Computational analysis of peripheral blood RNA sequencing data unravels disrupted immune patterns in Alzheimer's disease[J]. AIMS Neuroscience, 2024, 11(2): 103-117. doi: 10.3934/Neuroscience.2024007
The central nervous system (CNS) and the immune system collectively coordinate cellular functionalities, sharing common developmental mechanisms. Immunity-related molecules exert an influence on brain development, challenging the conventional view of the brain as immune-privileged. Chronic inflammation emerges as a key player in the pathophysiology of Alzheimer's disease (AD), with increased stress contributing to the disease progression and potentially exacerbating existing symptoms. In this study, the most significant gene signatures from selected RNA-sequencing (RNA-seq) data from AD patients and healthy individuals were obtained and a functional analysis and biological interpretation was conducted, including network and pathway enrichment analysis. Important evidence was reported, such as enrichment in immune system responses and antigen processes, as well as positive regulation of T-cell mediated cytotoxicity and endogenous and exogenous peptide antigen, thus indicating neuroinflammation and immune response participation in disease progression. These findings suggest a disturbance in the immune infiltration of the peripheral immune environment, providing new challenges to explore key biological processes from a molecular perspective that strongly participate in AD development.

| [1] |
DeTure MA, Dickson DW (2019) The neuropathological diagnosis of Alzheimer's disease. Mol Neurodegener 14: 1-18. https://doi.org/10.1186/s13024-019-0333-5 
|
| [2] |
Petersen RC, Lopez O, Armstrong MJ, et al. (2018) Practice guideline update summary: Mild cognitive impairment: Report of the Guideline Development, Dissemination, and Implementation Subcommittee of the American Academy of Neurology. Neurology 90: 126-135. https://doi.org/10.1212/WNL.0000000000004826
|
| [3] |
Breijyeh Z, Karaman R (2020) Comprehensive review on Alzheimer's disease: Causes and treatment. Molecules 25: 5789. https://doi.org/10.3390/molecules25245789
|
| [4] |
Golovatscka V, Ennes H, Mayer EA, et al. (2012) Chronic stress-induced changes in pro-inflammatory cytokines and spinal glia markers in the rat: a time course study. Neuroimmunomodulation 19: 367-376. https://doi.org/10.1159/000342092
|
| [5] |
Sheth C, McGlade E, Yurgelun-Todd D (2017) Chronic stress in adolescents and its neurobiological and psychopathological consequences: an RDoC perspective. Chronic Stress 1: 2470547017715645. https://doi.org/10.1177/2470547017715645
|
| [6] |
Lananna BV, Musiek ES (2020) The wrinkling of time: Aging, inflammation, oxidative stress, and the circadian clock in neurodegeneration. Neurobiol Dis 139: 104832. https://doi.org/10.1016/j.nbd.2020.104832
|
| [7] | Du X, Pang TY (2015) Is dysregulation of the HPA-axis a core pathophysiology mediating co-morbid depression in neurodegenerative diseases?. Front Psychiatry 6: 32. https://doi.org/10.3389/fpsyt.2015.00032 |
| [8] |
Sung PS, Lin PY, Liu CH, et al. (2020) Neuroinflammation and neurogenesis in Alzheimer's disease and potential therapeutic approaches. Int J Mol Sci 21: 7018. https://doi.org/10.3390/ijms21030701
|
| [9] |
Morimoto K, Nakajima K (2019) Role of the immune system in the development of the central nervous system. Front Neurosci 13: 916. https://doi.org/10.3389/fnins.2019.00916
|
| [10] |
Castellani G, Schwartz M (2020) Immunological features of non-neuronal brain cells: implications for Alzheimer's disease immunotherapy. Trends Immunol 41: 794-804. https://doi.org/10.1016/j.it.2020.07.005
|
| [11] |
Gagliardi S, Truffi M, Tinelli V, et al. (2022) Bisdemethoxycurcumin (BDC)-loaded H-ferritin-nanocages mediate the regulation of inflammation in Alzheimer's disease patients. Int J Mol Sci 23: 9237. https://doi.org/10.3390/ijms23169237
|
| [12] |
Prieto C, Barrios D (2020) RaNA-Seq: Interactive RNA-Seq analysis from FASTQ files to functional analysis. Bioinformatics 36: 1955-1956. https://doi.org/10.1093/bioinformatics/btz854
|
| [13] |
Schulze SK, Kanwar R, Gölzenleuchter M, et al. (2012) SERE: single-parameter quality control and sample comparison for RNA-Seq. BMC Genomics 13: 1-9. https://doi.org/10.1186/1471-2164-13-524
|
| [14] |
Patro R, Duggal G, Love MI, et al. (2017) Salmon provides fast and bias-aware quantification of transcript expression. Nat Methods 14: 417-419. https://doi.org/10.1038/nmeth.4197
|
| [15] |
Love MI, Huber W, Anders S (2014) Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 15: 1-21. https://doi.org/10.1186/s13059-014-0550-8
|
| [16] |
Szklarczyk D, Gable AL, Lyon D, et al. (2019) STRING v11: protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res 47: D607-D613. https://doi.org/10.1093/nar/gky1131
|
| [17] |
Chen EY, Tan CM, Kou Y, et al. (2013) Enrichr: interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinformatics 14: 1-14. https://doi.org/10.1186/1471-2105-14-128
|
| [18] |
Garcia-Moreno A, López-Domínguez R, Villatoro-García JA, et al. (2022) Functional enrichment analysis of regulatory elements. Biomedicines 10: 590. https://doi.org/10.3390/biomedicines10030590
|
| [19] |
Shannon P, Markiel A, Ozier O, et al. (2003) Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res 13: 2498-2504. https://doi.org/10.1101/gr.1239303
|
| [20] |
Gleimer M, Parham P (2003) Stress management: MHC class I and class I-like molecules as reporters of cellular stress. Immunity 19: 469-477. https://doi.org/10.1016/S1074-7613(03)00272-3
|
| [21] |
Wang BY, Ye YY, Qian C, et al. (2021) Stress increases MHC-I expression in dopaminergic neurons and induces autoimmune activation in Parkinson's disease. Neural Regen Res 16: 2521. https://doi.org/10.4103/1673-5374.313057
|
| [22] |
Candore G, Balistreri CR, Colonna-Romano G, et al. (2004) Major histocompatibility complex and sporadic Alzheimer's disease: a critical reappraisal. Exp Gerontol 39: 645-652. https://doi.org/10.1016/j.exger.2003.10.027
|
| [23] |
Gate D, Saligrama N, Leventhal O, et al. (2020) Clonally expanded CD8 T cells patrol the cerebrospinal fluid in Alzheimer's disease. Nature 577: 399-404. https://doi.org/10.1038/s41586-019-1895-7
|
| [24] |
Lomiguen C, Vidal L, Kozlowski P, et al. (2018) Possible role of chitin-like proteins in the etiology of Alzheimer's disease. J Alzheimers Disease 66: 439-444. https://doi.org/10.3233/JAD-180326
|
| [25] |
Russo C, Valle MS, Casabona A, et al. (2023) Chitinase Signature in the Plasticity of Neurodegenerative Diseases. Int J Mol Sci 24: 6301. https://doi.org/10.3390/ijms24076301
|
| [26] | Džoljić E, Grabatinić I, Kostić V (2015) Why is nitric oxide important for our brain?. Funct Neurol 30: 159. https://doi.org/10.11138/FNeur/2015.30.3.159 |
| [27] |
Huang WJ, Zhang XIA, Chen WW (2016) Role of oxidative stress in Alzheimer's disease. Biomed Rep 4: 519-522. https://doi.org/10.3892/br.2016.630
|
| [28] |
Skoumalová A, Hort J (2012) Blood markers of oxidative stress in A lzheimer's disease. J Cell Mol Med 16: 2291-2300. https://doi.org/10.1111/j.1582-4934.2012.01585.x
|
| [29] |
Kim M, Serwa RA, Samluk L, et al. (2023) Immunoproteasome-specific subunit PSMB9 induction is required to regulate cellular proteostasis upon mitochondrial dysfunction. Nat Commun 14: 4092. https://doi.org/10.1038/s41467-023-39642-8
|
| [30] |
Lorenzl S, Albers DS, Relkin N, et al. (2003) Increased plasma levels of matrix metalloproteinase-9 in patients with Alzheimer's disease. Neurochem Int 43: 191-19. https://doi.org/10.1016/S0197-0186(03)00004-4
|
| [31] |
Gu D, Liu F, Meng M, et al. (2020) Elevated matrix metalloproteinase-9 levels in neuronal extracellular vesicles in Alzheimer's disease. Ann Clin Transl Neur 7: 1681-1691. https://doi.org/10.1002/acn3.51155
|
| [32] |
Zipfel P, Rochais C, Baranger K, et al. (2020) Matrix metalloproteinases as new targets in Alzheimer's disease: Opportunities and challenges. J Med Chem 63: 10705-10725. https://doi.org/10.1021/acs.jmedchem.0c00352
|
| [33] |
Connolly K, Lehoux M, O'Rourke R, et al. (2023) Potential role of chitinase-3-like protein 1 (CHI3L1/YKL-40) in neurodegeneration and Alzheimer's disease. Alzh Dement 19: 9-24. https://doi.org/10.1002/alz.12612
|
| [34] |
Jorfi M, Maaser-Hecker A, Tanzi RE (2023) The neuroimmune axis of Alzheimer's disease. Genome Med 15: 1-2. https://doi.org/10.1186/s13073-023-01155-w
|
| [35] |
Lueg G, Gross CC, Lohmann H, et al. (2015) Clinical relevance of specific T-cell activation in the blood and cerebrospinal fluid of patients with mild Alzheimer's disease. Neurobiol Aging 36: 81-89. https://doi.org/10.1016/j.neurobiolaging.2014.08.008
|
| [36] |
Machado A, Herrera AJ, de Pablos RM, et al. (2014) Chronic stress as a risk factor for Alzheimer's disease. Rev Neuroscience 25: 785-804. https://doi.org/10.1515/revneuro-2014-0035
|
| [37] |
Olufunmilayo EO, Gerke-Duncan MB, Holsinger RD (2023) Oxidative Stress and Antioxidants in Neurodegenerative disorders. Antioxidants 12: 517. https://doi.org/10.3390/antiox12020517
|
| [38] |
Hampel H, Hardy J, Blennow K, et al. (2021) The amyloid-β pathway in Alzheimer's disease. Mol Psychiatr 26: 5481-5503. https://doi.org/10.1038/s41380-021-01249-0
|
| [39] |
Bettcher BM, Tansey MG, Dorothée G, et al. (2021) Peripheral and central immune system crosstalk in Alzheimer disease—a research prospectus. Nat Rev Neurol 17: 689-701. https://doi.org/10.1038/s41582-021-00549-x
|
| [40] |
Bettcher BM, Neuhaus J, Wynn MJ (2019) Increases in a pro-inflammatory chemokine, MCP-1, are related to decreases in memory over time. Front Aging Neurosci 11: 25. https://doi.org/10.3389/fnagi.2019.00025
|
| [41] |
Walker KA, Gross AL, Moghekar AR (2020) Association of peripheral inflammatory markers with connectivity in large-scale functional brain networks of non-demented older adults. Brain Behav Immun 87: 388-396. https://doi.org/10.1016/j.bbi.2020.01.006
|
| [42] |
Ouanes S, Popp J (2019) High cortisol and the risk of dementia and Alzheimer's disease: a review of the literature. Front Aging Neurosci 11: 43. https://doi.org/10.3389/fnagi.2019.00043
|
| [43] |
Ravi M, Miller AH, Michopoulos V (2021) The immunology of stress and the impact of inflammation on the brain and behaviour. BJPsych Adv 27: 158-165. https://doi.org/10.1192/bja.2020.82
|
| [44] |
Rothman SM, Mattson MP (2010) Adverse stress, hippocampal networks, and Alzheimer's disease. Neuromol Med 12: 56-70. https://doi.org/10.1007/s12017-009-8107-9
|
| [45] |
Krokidis MG, Vrahatis AG, Lazaros K, et al. (2023) Machine Learning Analysis of Alzheimer's Disease Single-Cell RNA-Sequencing Data across Cortex and Hippocampus Regions. Curr Issues Mol Biol 45: 8652-8669. https://doi.org/10.3390/cimb45110544
|
| [46] |
Ávila-Villanueva M, Gómez-Ramírez J, Maestú F, et al. (2020) The role of chronic stress as a trigger for the alzheimer disease continuum. Front Aging Neurosci 12: 561504. https://doi.org/10.3389/fnagi.2020.561504
|
| [47] |
Li Y, Qin J, Yan J, et al. (2019) Differences of physical vs. psychological stress: evidences from glucocorticoid receptor expression, hippocampal subfields injury, and behavioral abnormalities. Brain Imaging Behav 13: 1780-1788. https://doi.org/10.1007/s11682-018-9956-3
|
| [48] |
Thome AD, Faridar A, Beers DR, et al. (2018) Functional alterations of myeloid cells during the course of Alzheimer's disease. Mol Neurodegener 13: 1-11. https://doi.org/10.1186/s13024-018-0293-1
|
| [49] |
Lu Y, Li K, Hu Y, et al. (2021) Expression of Immune Related Genes and Possible Regulatory Mechanisms in Alzheimer's Disease. Front Immunol 12: 768966. https://doi.org/10.3389/fimmu.2021.768966
|
| [50] |
Wu Y, Liang S, Zhu H, et al. (2021) Analysis of immune-related key genes in Alzheimer's disease. Bioengineered 12: 9610-9624. https://doi.org/10.1080/21655979.2021.1999553
|
| [51] |
Ringland C, Schweig JE, Eisenbaum M, et al. (2021) MMP9 modulation improves specific neurobehavioral deficits in a mouse model of Alzheimer's disease. BMC Neurosci 22: 39. https://doi.org/10.1186/s12868-021-00643-2
|
| [52] |
Orre M, Kamphuis W, Dooves S, et al. (2013) Reactive glia show increased immunoproteasome activity in Alzheimer's disease. Brain 136: 1415-1431. https://doi.org/10.1093/brain/awt083
|
| [53] |
Mishto M, Bellavista E, Santoro A, et al. (2006) Immunoproteasome and LMP2 polymorphism in aged and Alzheimer's disease brains. Neurobiol Aging 27: 54-66. https://doi.org/10.1016/j.neurobiolaging.2004.12.004
|
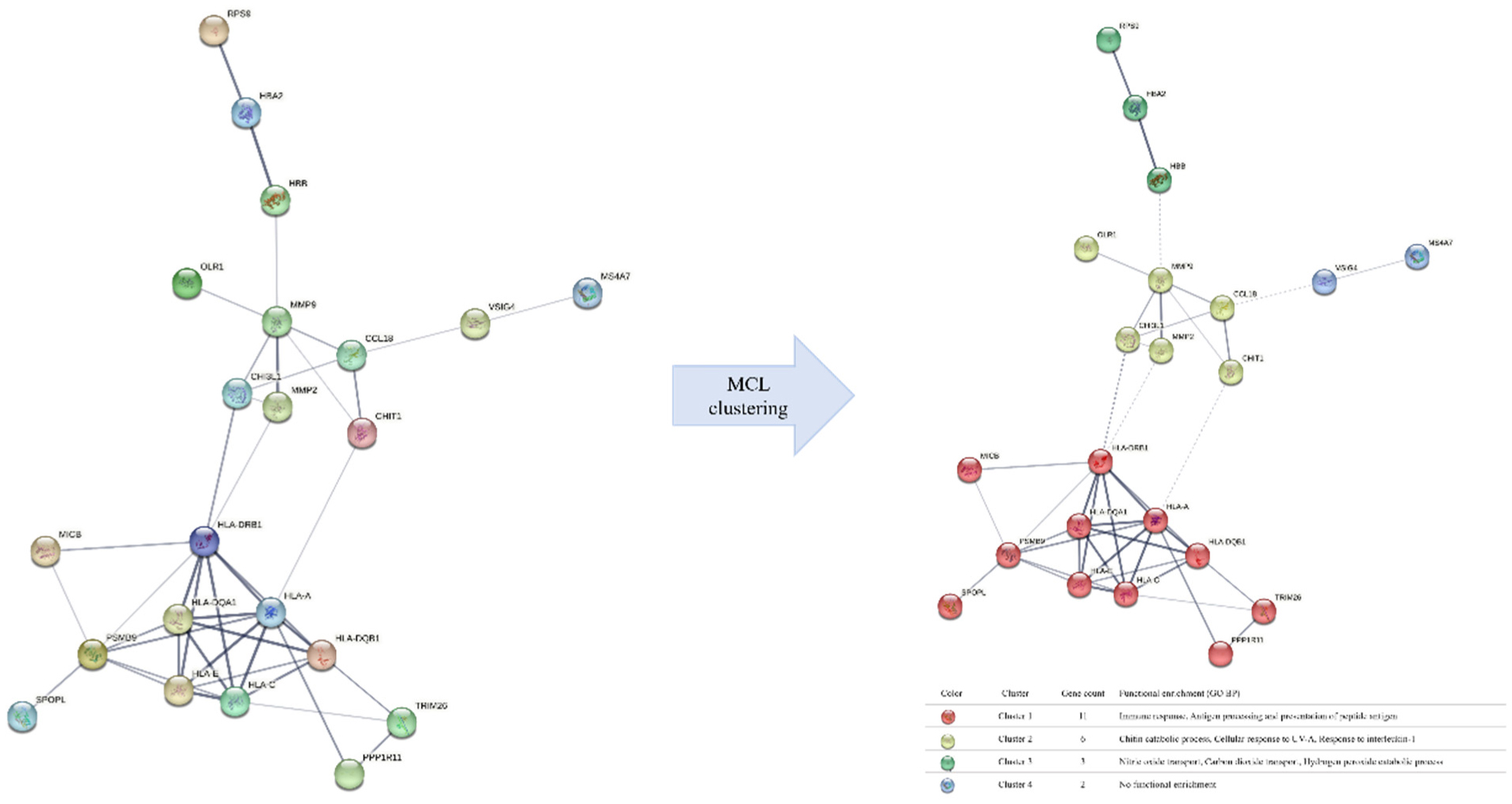
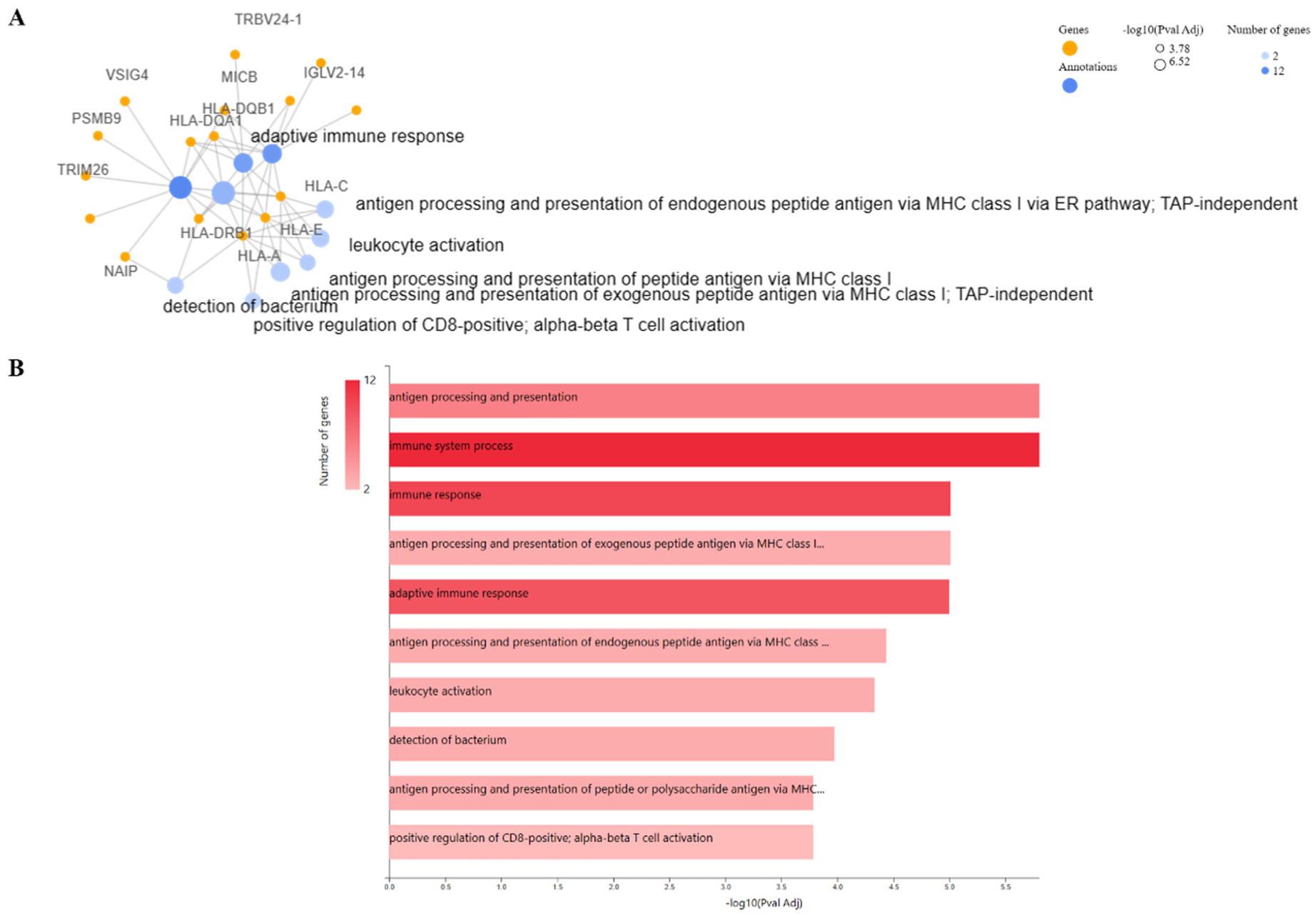
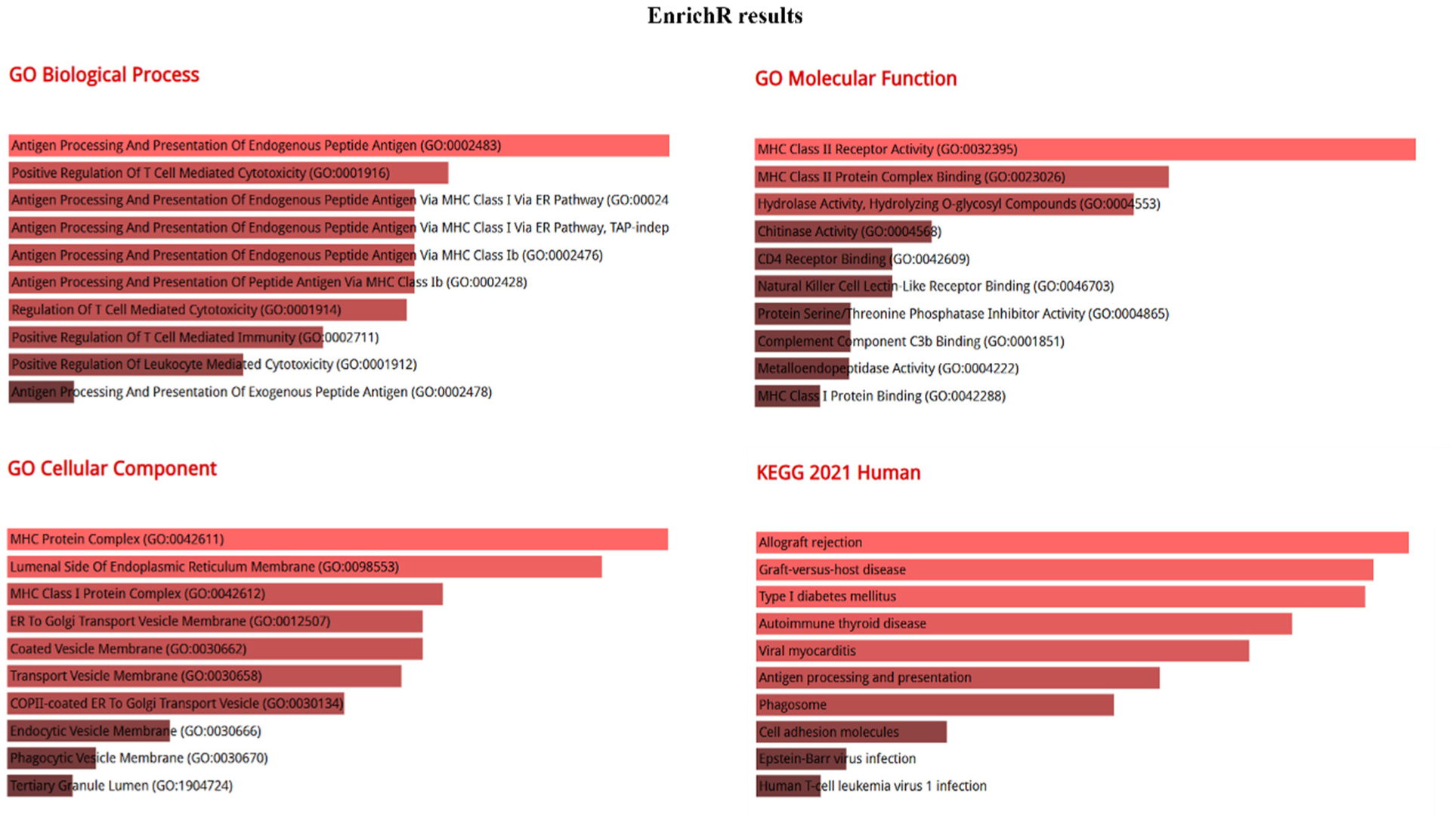
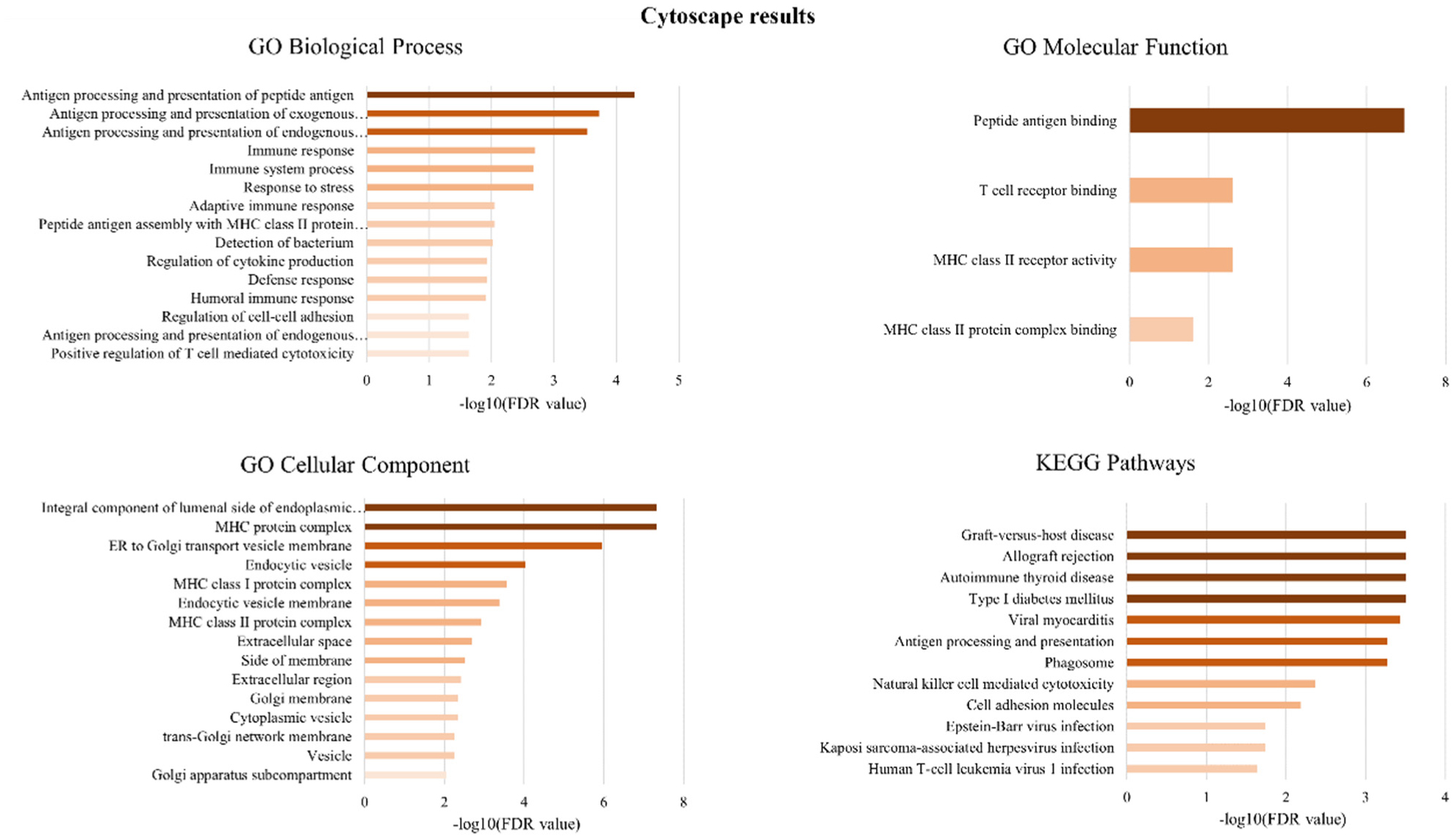
Figures(4) / Tables(1)

Dimitra Anatolou, Marios G. Krokidis. Computational analysis of peripheral blood RNA sequencing data unravels disrupted immune patterns in Alzheimer's disease[J]. AIMS Neuroscience, 2024, 11(2): 103-117. doi: 10.3934/Neuroscience.2024007







 DownLoad:
DownLoad: