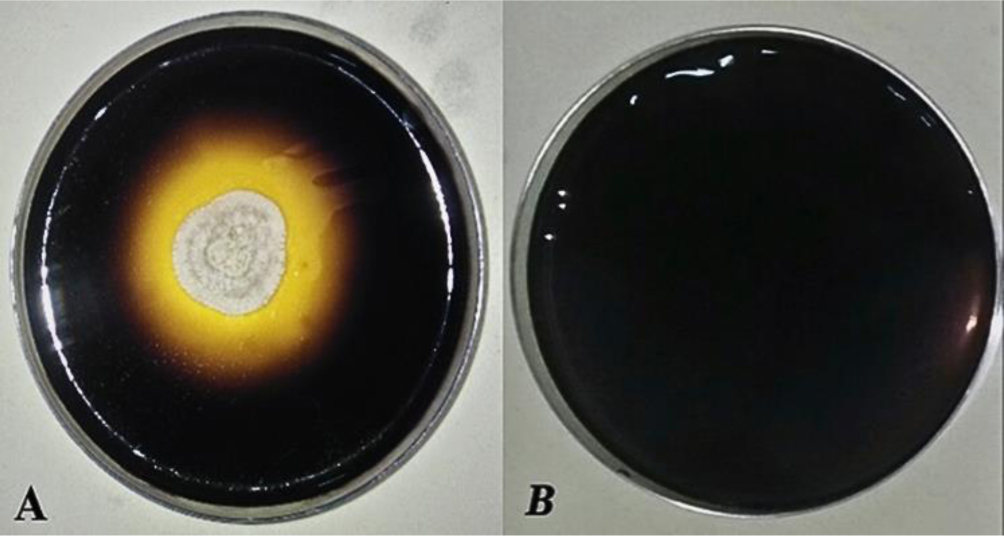
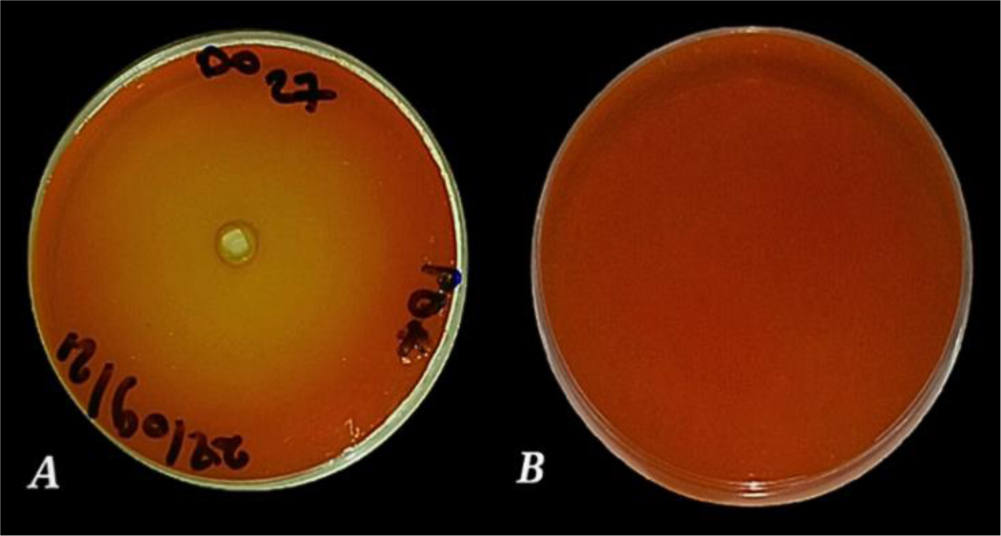
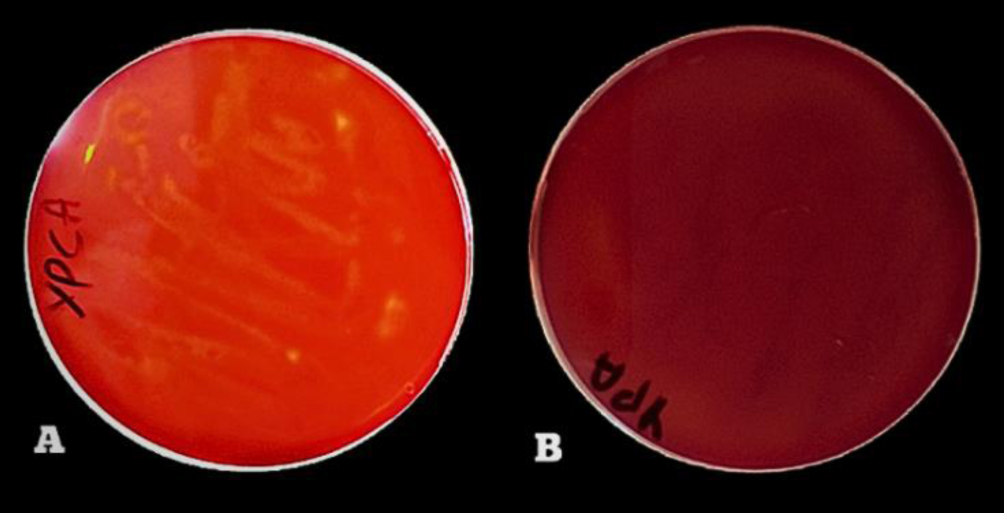
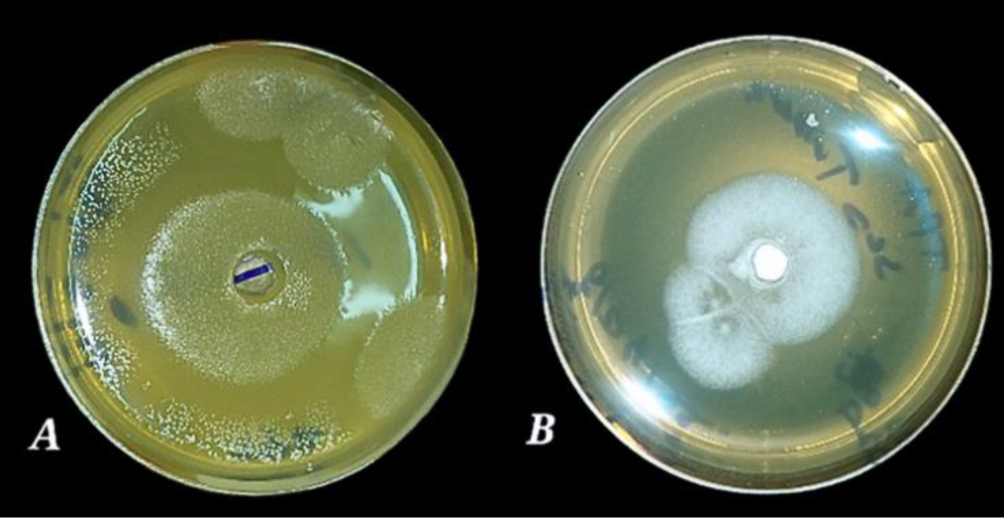
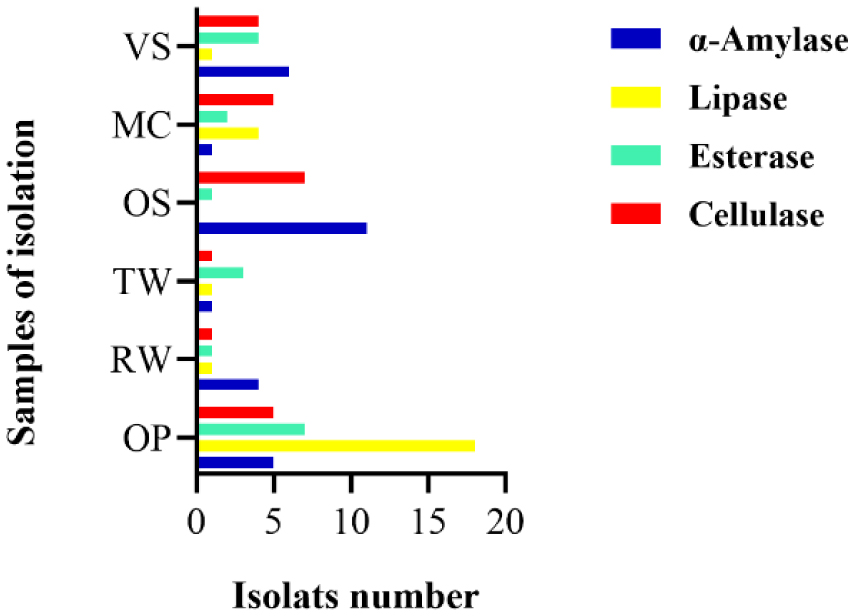
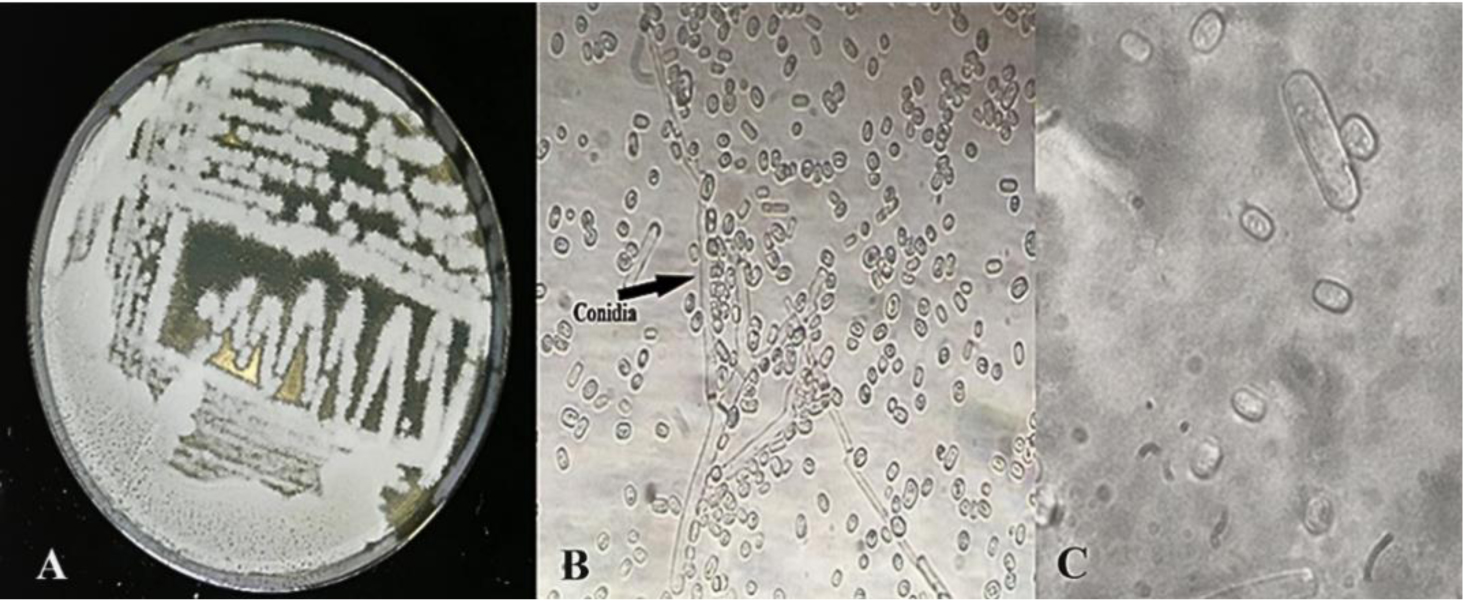
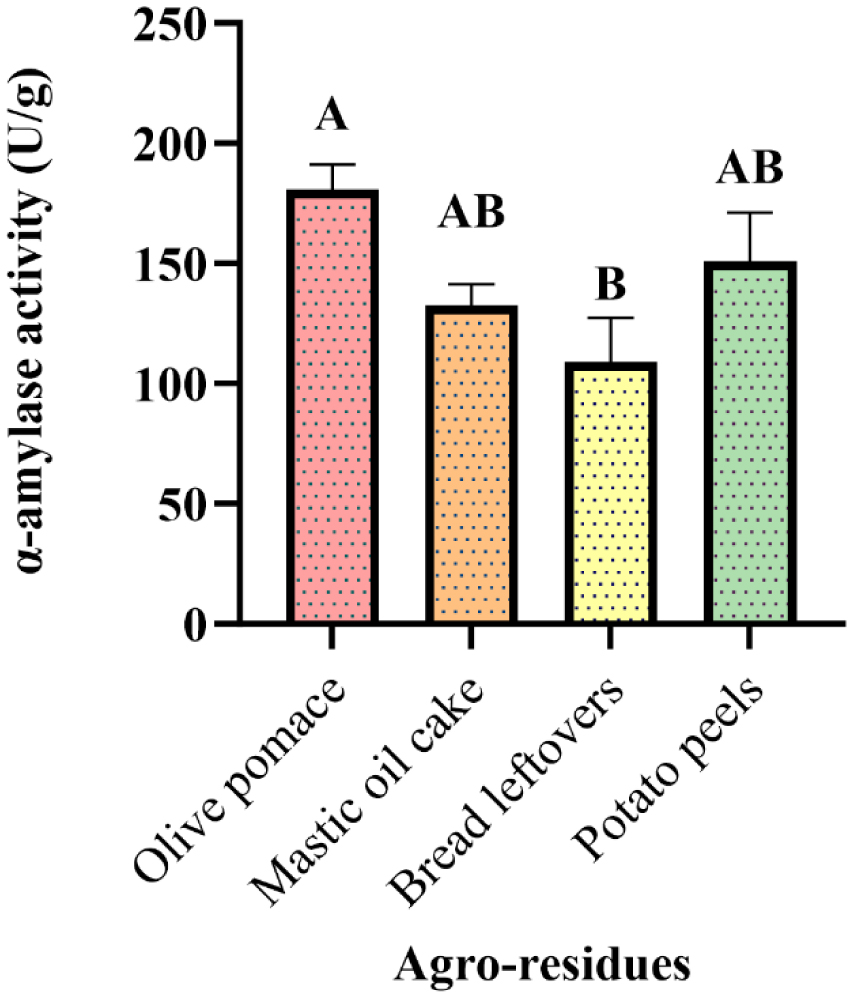
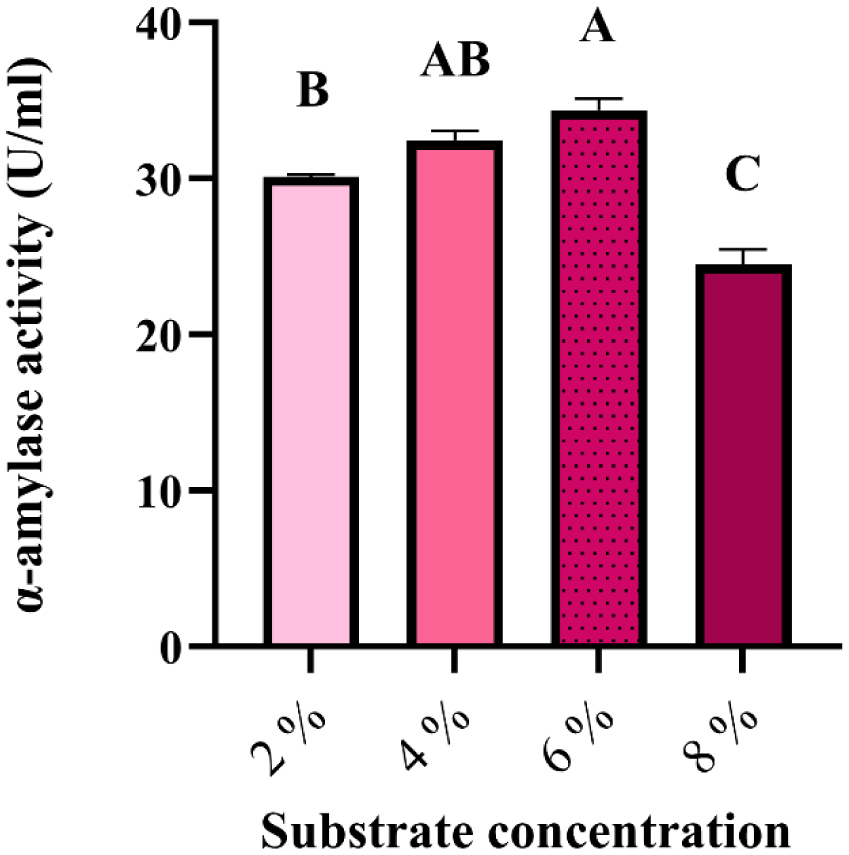
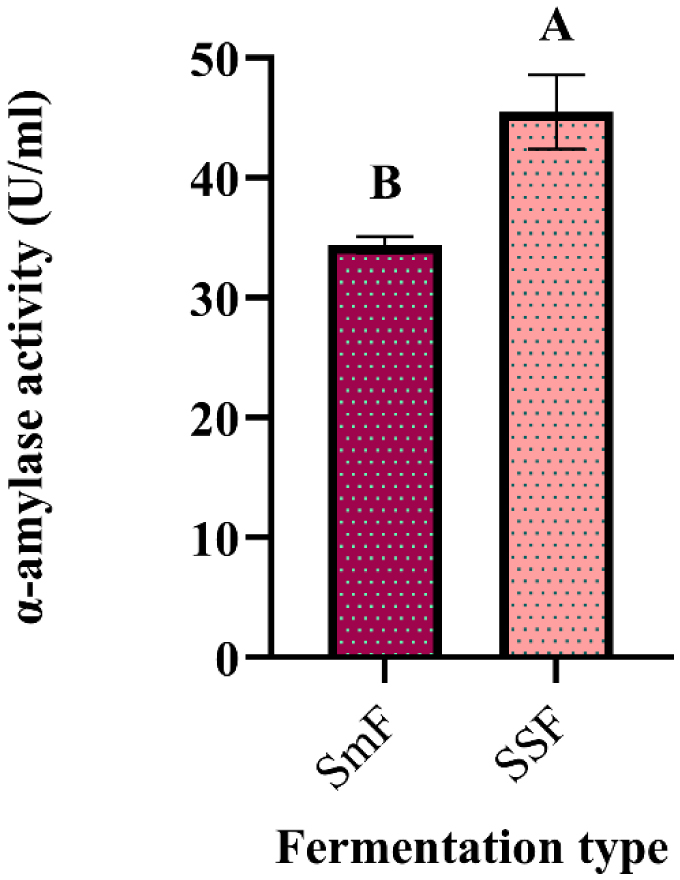
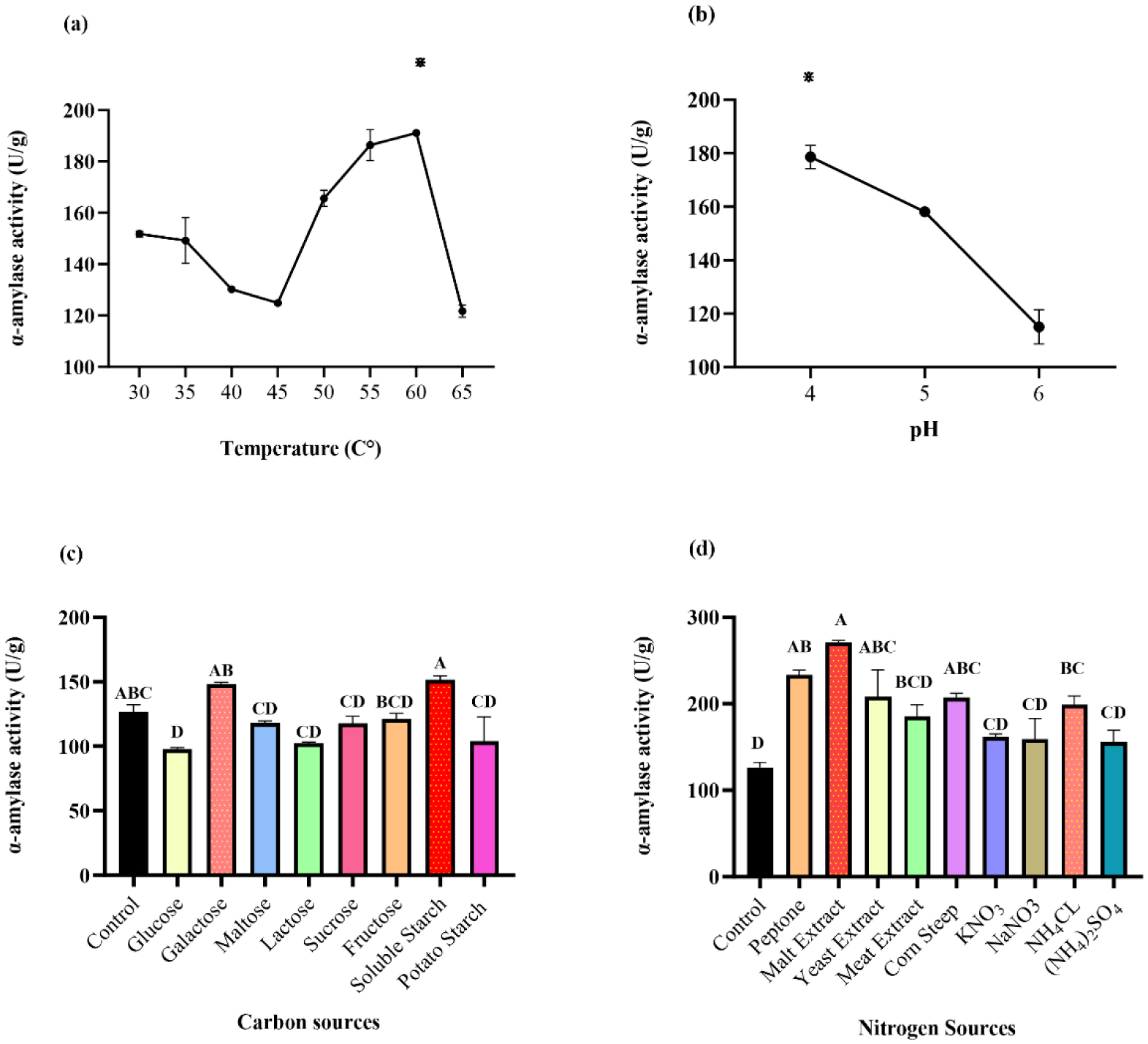
Enzymes are biocatalysts mainly used for their industrial potential in various applications. The present study aims to understand the enzyme production for biotechnological interest from a local yeast strain. From 100 isolates obtained from various biotopes, 78 strains were selected for their enzymatic heritage. Screening of α-amylase, lipase/esterase, and cellulase activities by rapid plate detection methods was carried out and the PO27 yeast was selected for its high capacity to produce α-amylase. In addition, this yeast strain exhibited good lipolytic and esterolytic activities, as well as low cellulase activity. A sequence analysis of the D1/D2 region of the 26S ribosomal RNA (26S rRNA) and a study of morphological characteristics identified the PO27 strain as Geotrichum candidum. The production of α-amylase has been studied in solid medium fermentation using various natural substrates without any supplementation such as olive pomace, potato peels, leftover bread, and mastic cake. G. candidum PO27 showed an improved production of α-amylase with olive pomace, thus reaching approximately 180.71 U/g. To evaluate the ability of this isolate to produce α-amylase in submerged fermentation, multiple concentrations of olive pomace substrate were tested. The best activity of submerged fermentation was statistically compared to the solid-state fermentation result in order to select the appropriate fermentation type. A high significant difference was found to rank the 6% olive pomace medium as the best substrate concentration with 34.395 U/mL of α-amylase activity. This work showed that the new isolate Geotrichum candidum PO27 has a better potential to produce α-amylase at a low cost in solid-state fermentation compared to submerged fermentation. Optimization conditions for PO27 α-amylase production through solid-state fermentation were achieved using a one factor at a time (OFAT) approach. The findings revealed that a high temperature (60 °C), an acidic pH, malt extract, and soluble starch were the highly significant medium components for enhancing α-amylase production. The use of olive pomace waste by Geotrichum candidum PO27 is expected to be effective in producing an industrially useful α-amylase.
Citation: Ibtissem Chaib, Scheherazed Dakhmouche-Djekrif, Leila Bennamoun, Tahar Nouadri. Extracellular enzymes producing yeasts study: cost-effective production of α-amylase by a newly isolated thermophilic yeast Geotrichum candidum PO27[J]. AIMS Microbiology, 2024, 10(1): 83-106. doi: 10.3934/microbiol.2024006
Enzymes are biocatalysts mainly used for their industrial potential in various applications. The present study aims to understand the enzyme production for biotechnological interest from a local yeast strain. From 100 isolates obtained from various biotopes, 78 strains were selected for their enzymatic heritage. Screening of α-amylase, lipase/esterase, and cellulase activities by rapid plate detection methods was carried out and the PO27 yeast was selected for its high capacity to produce α-amylase. In addition, this yeast strain exhibited good lipolytic and esterolytic activities, as well as low cellulase activity. A sequence analysis of the D1/D2 region of the 26S ribosomal RNA (26S rRNA) and a study of morphological characteristics identified the PO27 strain as Geotrichum candidum. The production of α-amylase has been studied in solid medium fermentation using various natural substrates without any supplementation such as olive pomace, potato peels, leftover bread, and mastic cake. G. candidum PO27 showed an improved production of α-amylase with olive pomace, thus reaching approximately 180.71 U/g. To evaluate the ability of this isolate to produce α-amylase in submerged fermentation, multiple concentrations of olive pomace substrate were tested. The best activity of submerged fermentation was statistically compared to the solid-state fermentation result in order to select the appropriate fermentation type. A high significant difference was found to rank the 6% olive pomace medium as the best substrate concentration with 34.395 U/mL of α-amylase activity. This work showed that the new isolate Geotrichum candidum PO27 has a better potential to produce α-amylase at a low cost in solid-state fermentation compared to submerged fermentation. Optimization conditions for PO27 α-amylase production through solid-state fermentation were achieved using a one factor at a time (OFAT) approach. The findings revealed that a high temperature (60 °C), an acidic pH, malt extract, and soluble starch were the highly significant medium components for enhancing α-amylase production. The use of olive pomace waste by Geotrichum candidum PO27 is expected to be effective in producing an industrially useful α-amylase.
submerged fermentation
state fermentation
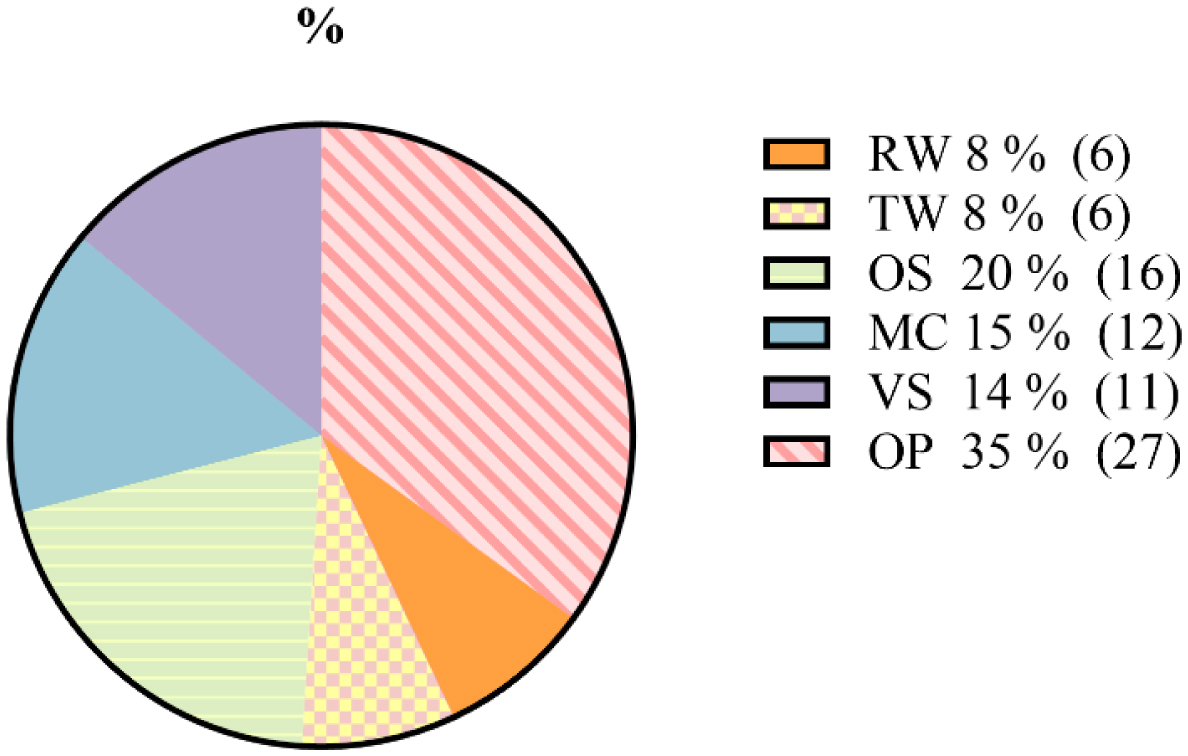
olive pomace
olive forest soil
dinitro salicylic acid
thermal water
mastic oil cake
vegetable smen
olive rinse water
yeast extract peptone soluble starch agar
yeast extract peptone dextrose agar
yeast extract malt extract
sabouraud dextrose agar
yeast extract glucose agar
yeast extract peptone carboxymethyl cellulose agar
tween peptone agar
amylolytic index
one factor at a time
National Center for Biotechnology Information

| [1] |
Pellis A, Cantone S, Ebert C, et al. (2018) Evolving biocatalysis to meet bioeconomy challenges and opportunities. New Biotechnol 40: 154-169. https://doi.org/10.1016/j.nbt.2017.07.005 
|
| [2] |
Costa-Silva TA, Carvalho AKF, Souza CRF, et al. (2021) Enhancement lipase activity via immobilization onto chitosan beads used as seed particles during fluidized bed drying: Application in butyl butyrate production. Appl Catal Gen 622: 118217. https://doi.org/10.1016/j.apcata.2021.118217
|
| [3] |
Okpara MO (2022) Microbial enzymes and their applications in food industry: A mini-review. Adv Enzyme Res 10: 23-47. https://doi.org/10.4236/aer.2022.101002
|
| [4] |
Vachher M, Sen A, Kapila R, et al. (2021) Microbial therapeutic enzymes: A promising area of biopharmaceuticals. Curr Res Biotechnol 3: 195-208. https://doi.org/10.1016/j.crbiot.2021.05.006
|
| [5] |
Raina D, Kumar V, Saran S (2022) A critical review on exploitation of agro-industrial biomass as substrates for the therapeutic microbial enzymes production and implemented protein purification techniques. Chemosphere 294: 133712. https://doi.org/10.1016/j.chemosphere.2022.133712
|
| [6] |
Nigam P (2013) microbial enzymes with special characteristics for biotechnological applications. Biomolecules 3: 597-611. https://doi.org/10.3390/biom3030597
|
| [7] |
Dahiya D, Nigam PS (2021) An overview of three biocatalysts of pharmaceutical importance synthesized by microbial cultures. AIMS Microbiol 7: 124-137. https://doi.org/10.3934/microbiol.2021009
|
| [8] |
Trabelsi S, Ben Mabrouk S, Kriaa M, et al. (2019) The optimized production, purification, characterization, and application in the bread making industry of three acid-stable alpha-amylases isoforms from a new isolated Bacillus subtilis strain US586. J Food Biochem 43: e12826. https://doi.org/10.1111/jfbc.12826
|
| [9] | Ilesanmi OI, Adekunle AE, Omolaiye JA, et al. (2020) Isolation, optimization and molecular characterization of lipase producing bacteria from contaminated soil. Sci Afr 8: 00279. https://doi.org/10.1016/j.sciaf.2020.e00279 |
| [10] | Khadka S, Khadka D, Poudel RC, et al. (2022) production optimization and biochemical characterization of cellulase from Geobacillus sp. KP43 Isolated from hot spring water of Nepal. BioMed Res Int 2022: 1-12. https://doi.org/10.1155/2022/6840409 |
| [11] |
Kuncharoen N, Techo S, Savarajara A, et al. (2020) Identification and lipolytic activity of yeasts isolated from foods and wastes. Mycology 11: 279-286. https://doi.org/10.1080/21501203.2020.1745922
|
| [12] |
Zaky AS, Greetham D, Louis EJ, et al. (2016) A new isolation and evaluation method for marine-derived yeast spp. with potential applications in industrial biotechnology. J Microbiol Biotechnol 26: 1891-1907. https://doi.org/10.4014/jmb.1605.05074
|
| [13] |
Kanmani P, Kumaresan K, Aravind J (2015) Utilization of coconut oil mill waste as a substrate for optimized lipase production, oil biodegradation and enzyme purification studies in Staphylococcus pasteuri. Electron J Biotechnol 18: 20-28. https://doi.org/10.1016/j.ejbt.2014.11.003
|
| [14] |
Salgado V, Fonseca C, Lopes da Silva T, et al. (2020) Isolation and identification of Magnusiomyces capitatus as a lipase-producing yeast from olive mill wastewater. Waste Biomass Valorization 11: 3207-3221. https://doi.org/10.1007/s12649-019-00725-7
|
| [15] |
Vyas S, Chhabra M (2017) Isolation, identification and characterization of Cystobasidium oligophagum JRC1: A cellulase and lipase producing oleaginous yeast. Bioresour Technol 223: 250-258. https://doi.org/10.1016/j.biortech.2016.10.039
|
| [16] |
Ramachandran S, Patel AK, Nampoothiri KM, et al. (2004) Alpha amylase from a fungal culture grown on oil cakes and its properties. Braz Arch Biol Technol 47: 309-317. https://doi.org/10.1590/S1516-89132004000200019
|
| [17] |
Benabda O, M'hir S,Kasmi M, et al. (2019) Optimization of protease and amylase production by Rhizopus oryzae cultivated on bread waste using solid-state fermentation. J Chem 2019: 3738181. https://doi.org/10.1155/2019/3738181
|
| [18] |
Olakusehin VO, Oyedeji O (2022) Production of α-amylase from Aspergillus flavus S2-OY using solid substrate fermentation of potato (Solanum tuberosum L.) peel. Int J Biol Chem Sci 15: 1950-1967. https://doi.org/10.4314/ijbcs.v15i5.21
|
| [19] |
Salah HA, Temerk HA, Salah NA, et al. (2021) Production and optimization of xylanase and α-amylase from non-saccharomyces yeasts (Pichia membranifaciens). J Pure Appl Microbiol 15: 452-461. https://doi.org/10.22207/JPAM.15.1.43
|
| [20] |
Lee YJ, Choi YR, Lee SY, et al. (2011) Screening wild yeast strains for alcohol fermentation from various fruits. Mycobiology 39: 33-39. https://doi.org/10.4489/MYCO.2011.39.1.033
|
| [21] |
Buzzini P, Martini A (2002) Extracellular enzymatic activity profiles in yeast and yeast-like strains isolated from tropical environments. J Appl Microbiol 93: 1020-1025. https://doi.org/10.1046/j.1365-2672.2002.01783.x
|
| [22] | Riyadi FA, Alam Z, Salleh N, et al. (2017) Thermostable and organic solvent tolerant lipase producing fungi in solid state bioconversion of palm kernel cake. Asia Pac J Mol Biol Biotechnol 25: 98-105. |
| [23] | Lanka S, B TT (2018) Screening and isolation of lipase producing fungi from marine water obtained from Machilipatnam costal region. Int J Pharmacogn Phytochem Res 9: 7. https://doi.org/10.25258/phyto.v9i07.11157 |
| [24] |
Singh R, Gupta N, Goswami VK, et al. (2006) A simple activity staining protocol for lipases and esterases. Appl Microbiol Biotechnol 70: 679-682. https://doi.org/10.1007/s00253-005-0138-z
|
| [25] |
Carrim AJI, Barbosa EC, Vieira JDG (2006) Enzymatic activity of endophytic bacterial isolates of Jacaranda decurrens Cham. (Carobinha-do-campo). Braz Arch Biol Technol 49: 353-359. https://doi.org/10.1590/S1516-89132006000400001
|
| [26] |
Sacristán N, González L, Castro JM, et al. (2012) Technological characterization of Geotrichum candidum strains isolated from a traditional Spanish goats' milk cheese. Food Microbiol 30: 260-266. https://doi.org/10.1016/j.fm.2011.10.003
|
| [27] |
Sampaio JP, Gadanho M, Santos S, et al. (2001) Polyphasic taxonomy of the basidiomycetous yeast genus rhodosporidium: Rhodosporidium kratochvilovae and related anamorphic species. Int J Syst Evol Microbiol 51: 687-697. https://doi.org/10.1099/00207713-51-2-687
|
| [28] |
Turchetti B, Goretti M, Branda E, et al. (2013) Influence of abiotic variables on culturable yeast diversity in two distinct Alpine glaciers. FEMS Microbiol Ecol 86: 27-340. https://doi.org/10.1111/1574-6941.12164
|
| [29] | Saitou N, Nei M (1987) The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol Biol Evol 4: 406-425. https://doi.org/10.1093/oxfordjournals.molbev.a040454 |
| [30] |
Kimura M (1980) A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 16: 111-120. https://doi.org/10.1007/BF01731581
|
| [31] |
Dakhmouche Djekrif S, Bennamoun L, Labbani FZK, et al. (2021) An alkalothermophilic amylopullulanase from the yeast Clavispora lusitaniae ABS7: Purification, characterization and potential application in laundry detergent. Catalysts 11: 1438. https://doi.org/10.3390/catal11121438
|
| [32] |
Sahoo RK, Sahu A, Subudhi E (2020) Bioremediation of hydrocarbon using bacterial lipase from waste biomass. Iran J Sci Technol Trans Sci 44: 1287-1293. https://doi.org/10.1007/s40995-020-00935-y
|
| [33] |
Bernfeld P (1955) Amylases, α and β. Meth Enzymol 1: 149-158. https://doi.org/10.1016/0076-6879(55)01021-5
|
| [34] |
Oussadi MI, Kitouni M (2015) Statistical optimization of cultural conditions of an halophilic alpha-amylase production by halophilic Streptomyces sp. grown on orange waste powder. Biocatal Agric Biotechnol 4: 685-693. https://doi.org/10.1016/j.bcab.2015.08.011
|
| [35] | Rosa C, Gábor P (2006) Biodiversity and ecophysiology of yeasts. The yeast handbook . Berlin; London: Springer 155-174. https://doi.org/10.1007/3-540-30985-3 |
| [36] |
Buzzini P, Turchetti B, Yurkov A (2018) Extremophilic yeasts: the toughest yeasts around?. Yeast 35: 487-497. https://doi.org/10.1002/yea.3314
|
| [37] | Misbah A, Aouine M, Raouan SE, et al. (2019) Microorganisms isolated from Moroccan olive-mill wastes: screening of their enzymatic activities for biotechnological use. Eur Sci J ESJ 15: 464-494. https://doi.org/10.19044/esj.2019.v15n30p464 |
| [38] |
Williams KF, Agwa OK, Abu GO, et al. (2021) Screening, characterization and identification of sophorolipid-producing yeast isolated from palm oil effluent polluted soil. J Adv Microbiol 21: 1-16. https://doi.org/10.9734/jamb/2021/v21i1230409
|
| [39] |
Singh P, Kashyap K, Singh S (2014) Isolation, purification and characterization of oil degrading bacteria from different oil cake samples. Int J Bioinforma Biol Sci 2: 189. https://doi.org/10.5958/2321-7111.2014.00016.X
|
| [40] |
Sudan SK, Kumar N, Kaur I, et al. (2018) Production, purification and characterization of raw starch hydrolyzing thermostable acidic α-amylase from hot springs, India. Int J Biol Macromol 117: 831-839. https://doi.org/10.1016/j.ijbiomac.2018.05.231
|
| [41] | Karam NE, Idoui T (2008) Lactic acid bacteria from Jijel's traditional butter: Isolation, Identification and major technological traits. Grasas y Aceites 59: 361-367. https://doi.org/10.3989/gya.2008.v59.i4.530 |
| [42] |
Bintsis T (2021) Yeasts in different types of cheese. AIMS Microbiol 7: 447-470. https://doi.org/10.3934/microbiol.2021027
|
| [43] |
Kwon YM, Choi HS, Lim JY, et al. (2020) Characterization of amylolytic activity by a marine-derived yeast Sporidiobolus pararoseus PH-Gra1. Mycobiology 48: 95-203. https://doi.org/10.1080/12298093.2020.1763100
|
| [44] |
Gunam IBW, Sujana IGA, Wijaya IMM, et al. (2021) Isolation and selection of amylase-producing microbes isolated from ragi tape and cassava tape available on the markets. IOP Conf Ser Earth Environ Sci 913: 012041. https://doi.org/10.1088/1755-1315/913/1/012041
|
| [45] | Muriithi J, Matofari J, Nduko J (2021) Amylolytic microorganisms from diverse tropical environments: Isolation, identification, and amylase production. Appl Res 1: 1-9. https://doi.org/10.1002/appl.202100007 |
| [46] |
Lanka S, Latha JNL (2015) A Short review on various screening methods to isolate potential lipase producers: lipases-the present and future enzymes of biotech industry. Int J Biol Chem 9: 207-219. https://doi.org/10.3923/ijbc.2015.207.219
|
| [47] |
Bharathi D, Rajalakshmi G (2019) Microbial lipases: An overview of screening, production and purification. Biocatal Agric Biotechnol 22: 101368. https://doi.org/10.1016/j.bcab.2019.101368
|
| [48] | Arman Z, Sondana GA, Fikriyyah NN, et al. (2020) Screening of amylolytic and cellulolytic yeast from Dendrobium spathilingue in Bali Botanical Garden, Indonesia. AIP Conf Proc 2242: 050013-1-050013-5. https://doi.org/10.1063/5.0007802 |
| [49] | Wadia T, Jain SK (2017) Isolation, screening and identification of lipase producing fungi from oil contaminated soil of Shani Mandir Ujjain. Int J Curr Microbiol Appl Sci 6: 872-1878. https://doi.org/10.20546/ijcmas.2017.607.223 |
| [50] |
Zaier H, Maktouf S, Roussos S, et al. (2021) Filamentous fungi isolated from Tunisian olive mill wastes: use of solid-state fermentation for enzyme production. Not Bot Horti Agrobot Cluj-Napoca 49: 12125. https://doi.org/10.15835/nbha49112125
|
| [51] |
Hlavsová K, Zarevúcka M, Wimmer Z, et al. (2009) Geotrichum candidum 4013: Extracellular lipase versus cell-bound lipase from the single strain. J Mol Catal B Enzym 61: 188-193. https://doi.org/10.1016/j.molcatb.2009.06.012
|
| [52] |
Attanayaka DPSTG, De Silva SNT, Nirosha SF, et al. (2009) Isolation of raw starch hydrolysing fungi and purification of α-amylase from Geotrichum candidum CMSS06. J Natl Sci Found Sri Lanka 37: 93. https://doi.org/10.4038/jnsfsr.v37i2.1064
|
| [53] |
Gad AM, Suleiman WB, El-Sheikh HH, et al. (2022) Characterization of cellulase from Geotrichum candidum Strain Gad1 approaching bioethanol production. Arab J Sci Eng 47: 6837-6850. https://doi.org/10.1007/s13369-021-06391-z
|
| [54] | Rodionova NA, Dubovaia NV, Eneĭskaia EV, et al. (2000) Purification and characteristic of endo-(1--4)-beta-xylanase from Geotrichum candidum 3C. Prikl Biokhim Mikrobiol 36: 535-540. |
| [55] |
Ahmed A, Ejaz U, Sohail M (2020) Pectinase production from immobilized and free cells of Geotrichum candidum AA15 in galacturonic acid and sugars containing medium. J. King Saud Univ 32: 952-954. https://doi.org/10.1016/j.jksus.2019.07.003
|
| [56] |
Muhammad A, Bokhari SAI, Vernoux JP, et al. (2019) Purification, characterization and thermodynamic assessment of an alkaline protease by Geotrichum candidum of dairy origin. Iran J Biotechnol 17: e2042. https://doi.org/10.21859/ijb.2042
|
| [57] | Loo JL, Lai OM, Long K, et al. (2006) Identification and characterisation of a locally isolated lipolytic microfungus - Geotrichum candidum. Malays J Microbiol 2: 22-23. https://doi.org/10.21161/mjm.210604 |
| [58] |
Karam EA, Abdel Wahab WA, Saleh SAA, et al. (2017) Production, immobilization and thermodynamic studies of free and immobilized Aspergillus awamori amylase. Int J Biol Macromol 102: 694-703. https://doi.org/10.1016/j.ijbiomac.2017.04.033
|
| [59] |
Obi CN, Okezie O, Ezugwu AN (2019) Amylase Production by solid state fermentation of agro-industrial wastes using Bacillus species. Eur J Nutr Food Saf 9: 408-414. https://doi.org/10.9734/ejnfs/2019/v9i430087
|
| [60] |
Balakrishnan M, Jeevarathinam G, Kumar SKS, et al. (2021) Optimization and scale-up of α-amylase production by Aspergillus oryzae using solid-state fermentation of edible oil cakes. BMC Biotechnol 21: 33. https://doi.org/10.1186/s12896-021-00686-7
|
| [61] |
Khelil O, Choubane S, Maredj K, et al. (2022) UV mutagenesis for the overproduction of thermoalkali-stable α-amylase from Bacillus subtilis TLO3 by fermentation of stale bread: Potential application as detergent additive. Biocatal Agric Biotechnol 43: 02403. https://doi.org/10.1016/j.bcab.2022.102403
|
| [62] |
Benabda O, M'hir S, Kasmi M, et al. (2019) Optimization of protease and amylase production by Rhizopus oryzae cultivated on bread waste using solid-state fermentation. J Chem 2019: 1-9. https://doi.org/10.1155/2019/3738181
|
| [63] |
Pranay K, Padmadeo SR, Prasad B (2019) Production of amylase from Bacillus subtilis sp. strain KR1 under solid state fermentation on different agrowastes. Biocatal Agric Biotechnol 21: 101300. https://doi.org/10.1016/j.bcab.2019.101300
|
| [64] |
Almanaa TN, Vijayaraghavan P, Alharbi NS, et al. (2020) Solid state fermentation of amylase production from Bacillus subtilis D19 using agro-residues. J King Saud Univ - Sci 32: 1555-1561. https://doi.org/10.1016/j.jksus.2019.12.011
|
| [65] |
Djekrif-Dakhmouche S, Gheribi-Aoulmi Z, Meraihi Z, et al. (2006) Application of a statistical design to the optimization of culture medium for α-amylase production by Aspergillus niger ATCC 16404 grown on orange waste powder. J Food Eng 73: 190-197. https://doi.org/10.1016/j.jfoodeng.2005.01.021
|
| [66] |
Singh R, Langyan S, Sangwan S, et al. (2022) Optimization and production of alpha-amylase using Bacillus subtilis from apple peel: Comparison with alternate feedstock. Food Biosci 49: 101978. https://doi.org/10.1016/j.fbio.2022.101978
|
| [67] | Abdel-Mageed HM, Barakat AZ, Bassuiny RI, et al. (2022) Biotechnology approach using watermelon rind for optimization of α-amylase enzyme production from Trichoderma virens using response surface methodology under solid-state fermentation. Folia Microbiol (Praha) 67: 53-264. https://doi.org/10.1007/s12223-021-00929-2 |
| [68] | Divya K, Padma PN (2014) Yeast isolates from diverse sources for cold-active polygalacturonase and amylase production. Int J Sci Technol Res 3: 145-148. |
| [69] |
Falih AM (1998) Effect of heavy-metals on amylolytic activity of the soil yeasts Geotrichum capitatum and Geotrichum candidum. Bioresour Technol 66: 213-217. https://doi.org/10.1016/S0960-8524(98)00063-7
|
| [70] |
Najmalddin H, Yurdugül S, Hamzah H (2023) Screening of enzyme activities for improvement of bread quality by potato peel addition to the yeast growth medium. Food Biosci 51: 102239. https://doi.org/10.1016/j.fbio.2022.102239
|
| [71] |
Arunsasi M, Kani SM, Jegadeesh G, et al. (2010) Submerged fermentation of amylase enzyme by Aspergillus flavus using Cocos nucifera meal. Kathmandu Univ J Sci Eng Technol 6: 75-87. https://doi.org/10.3126/kuset.v6i2.4015
|
| [72] |
Jesubunmi C, Ogbonna J (2022) Production of cellulase and amylase enzymes in both solid and liquid states by two species of fungi. Asian J Biotechnol Bioresour Technol 8: 1-11. https://doi.org/10.9734/ajb2t/2022/v8i130115
|
| [73] |
Premalatha A, Vijayalakshmi K, Shanmugavel M, et al. (2022) Optimization of culture conditions for enhanced production of extracellular α-amylase using solid-state and submerged fermentation from Aspergillus tamarii MTCC5152. Biotechnol Appl Biochem 70: 835-845. https://doi.org/10.1002/bab.2403
|
| [74] |
Ahmed SA, Abdella MAA, El-Sherbiny GM, et al. (2019) Application of one–factor- at-a-time and statistical designs to enhance α-amylase production by a newly isolate Bacillus subtilis strain-MK1. Biocatal Agric Biotechnol 22: 101397. https://doi.org/10.1016/j.bcab.2019.101397
|
| [75] |
Luang-In V, Yotchaisarn M, Saengha W, et al. (2019) Isolation and identification of amylase-producing bacteria from soil in Nasinuan community forest, Maha Sarakham, Thailand. Biomed Pharmacol J 12: 1061-1068. https://doi.org/10.13005/bpj/1735
|
| [76] | Finore I, Paola Di Donato IF, Poli A, et al. (2014) Use of agro waste biomass for α-amylase production by anoxybacillus amylolyticus: purification and properties. J Microb Biochem Technol 06: 320-326. https://doi.org/10.4172/1948-5948.1000162 |
| [77] | Djekrif-Dakhmouche S (2016) Production and characterization of amylopullulanase from the yeast Clavispora lusitaniae abs7 isolated from wheat, grown and stored in arid zones, doctoral thesis. Compiegne University of Technology, (UTS), Compiegne France 211. |
| [78] | Mathew J, John N, Vazhacharickal P, et al. (2016) Comparative study of the activity of amylase produced by Aspergillus niger though solid state fermentation (SSF) using various starchy materials. Indian J Plant Sci 5: 2319-3824. |
| [79] | Tatsinkou Fossi B, Tavea F, Ndjouenkeu R (2005) Production and partial characterization of a thermostable amylase from ascomycetes yeast strain isolated from starchy soils. Afr J Biotechnol 4: 1684-5315. |
| [80] |
Olakusehin VO, Oyedeji O (2022) Production of α-amylase from Aspergillus flavus S2-OY using solid substrate fermentation of potato (Solanum tuberosum L.) peel. Int J Biol Chem Sci 15: 1950-1967. https://doi.org/10.4314/ijbcs.v15i5.21
|
| [81] |
Skaltsounis Al, Argyropoulou A, Aligiannis N, et al. (2015) Recovery of high added value compounds from olive tree products and olive processing by. Olive and Olive Oil Bioactive Constituents . AOCS Press 333-356. https://doi.org/10.1016/B978-1-63067-041-2.50017-3
|
| [82] |
De Mot R., Verachtert H. (1986) Secretion of alpha-amylase and multiple forms of glucoamylase by the yeast Trichosporon pullulans. Can J Microbiol 32: 47-51. https://doi.org/10.1139/m86-009
|
| [83] |
Al-Dhabi NA, Esmail GA, Ghilan AKM, et al. (2020) Isolation and purification of starch hydrolyzing amylase from Streptomyces sp. Al-Dhabi-46 obtained from the Jazan region of Saudi Arabia with industrial applications. J King Saud Univ 32: 1226-1232. https://doi.org/10.1016/j.jksus.2019.11.018
|
| [84] |
Ertan F, Balkan B, Balkan S, et al. (2006) Solid state fermentation for the production of α-amylase from Penicillium chrysogenum using mixed agricultural by-products as substrate. Biologia 61: 657-661. https://doi.org/10.2478/s11756-006-0137-2
|
| [85] |
Kizhakedathil MPJ, Subathra Devi C (2021) Acid stable α-amylase from Pseudomonas balearica VITPS19—Production, purification and characterization. Biotechnol Rep 30: e00603. https://doi.org/10.1016/j.btre.2021.e00603
|
| [86] |
Ahmed SA, Mostafa FA, Helmy WA, et al. (2017) Improvement of bacterial α-amylase production and application using two steps statistical factorial design. Biocatal Agric Biotechnol 10: 224-233. https://doi.org/10.1016/j.bcab.2017.03.004
|
 microbiol-10-01-006-s001.pdf microbiol-10-01-006-s001.pdf |
 |

Figures(12)

Ibtissem Chaib, Scheherazed Dakhmouche-Djekrif, Leila Bennamoun, Tahar Nouadri. Extracellular enzymes producing yeasts study: cost-effective production of α-amylase by a newly isolated thermophilic yeast Geotrichum candidum PO27[J]. AIMS Microbiology, 2024, 10(1): 83-106. doi: 10.3934/microbiol.2024006







 DownLoad:
DownLoad: