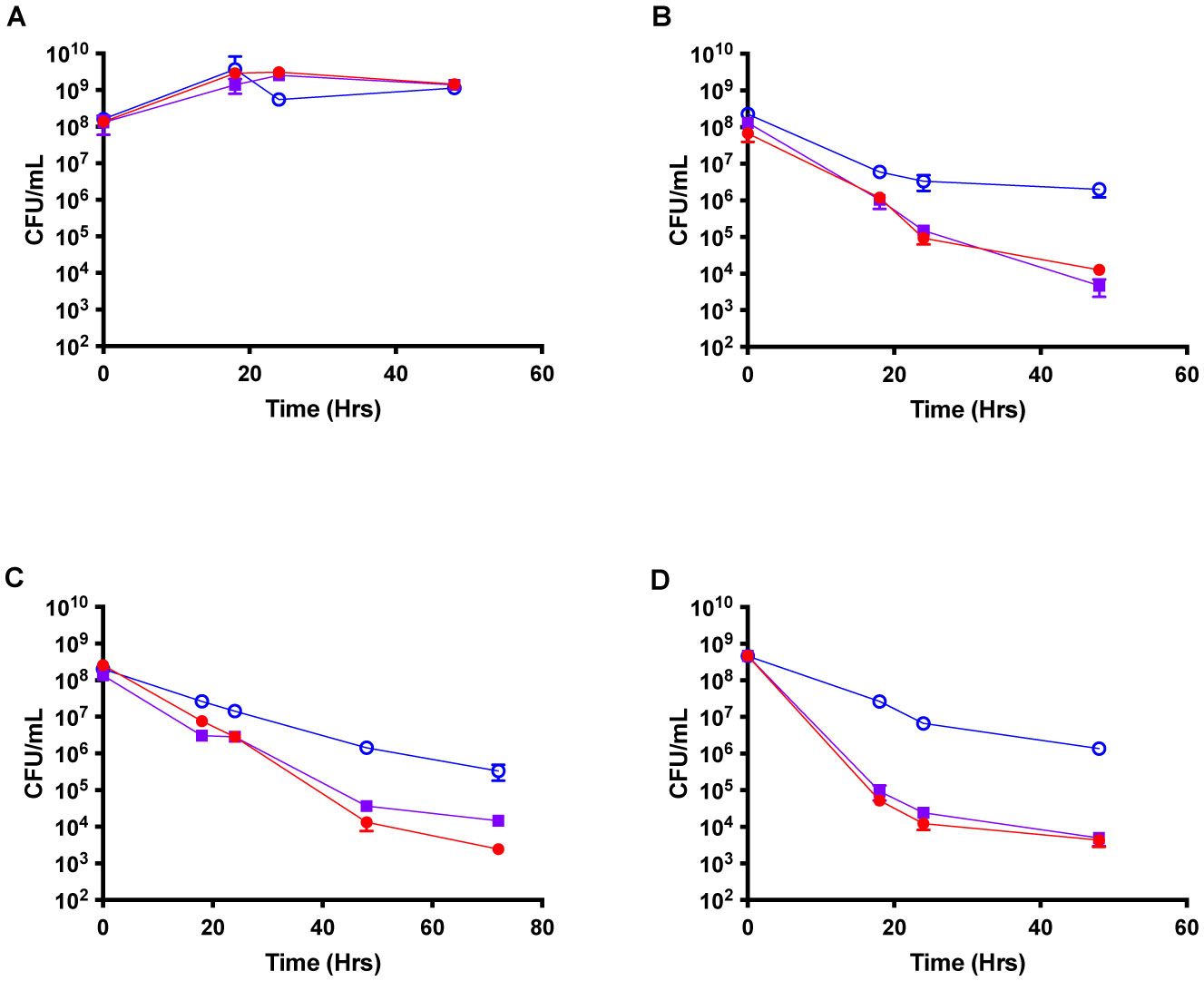
Staphylococcus aureus is widely known for its resistance and virulence causing public health concerns. However, antibiotic tolerance is also a contributor to chronic and relapsing infections. Previously, it has been demonstrated that persister formation is dependent on reduced tricarboxylic acid (TCA) cycle activity. Persisters have been extensively examined in terms of antibiotic tolerance but tolerance to antimicrobial peptides (AMPs) remains largely unexplored. AMPs are a key component of both the human and Drosophila innate immune response. TCA cycle mutants were tested to determine both antibiotic and AMP tolerance. Challenging with multiple classes of antibiotics led to increased persister formation (100- to 1,000-fold). Similarly, TCA mutants exhibited AMP tolerance with a 100- to 1,000-fold increase in persister formation when challenged with LL-37 or human β-defensin 3 (hβD3). The ability of TCA cycle mutants to tolerate the innate immune system was further examined with a D. melanogaster model. Both males and females infected with TCA cycle mutants exhibited increased mortality and had higher bacterial burdens (1.5 log) during the course of the infection. These results suggest increasing the percentage of persister cells leads to increased tolerance to components of the innate immune system.
Citation: Alexis M. Hobbs, Kennedy E. Kluthe, Kimberly A. Carlson, Austin S. Nuxoll. Interruption of the tricarboxylic acid cycle in Staphylococcus aureus leads to increased tolerance to innate immunity[J]. AIMS Microbiology, 2021, 7(4): 513-527. doi: 10.3934/microbiol.2021031
Staphylococcus aureus is widely known for its resistance and virulence causing public health concerns. However, antibiotic tolerance is also a contributor to chronic and relapsing infections. Previously, it has been demonstrated that persister formation is dependent on reduced tricarboxylic acid (TCA) cycle activity. Persisters have been extensively examined in terms of antibiotic tolerance but tolerance to antimicrobial peptides (AMPs) remains largely unexplored. AMPs are a key component of both the human and Drosophila innate immune response. TCA cycle mutants were tested to determine both antibiotic and AMP tolerance. Challenging with multiple classes of antibiotics led to increased persister formation (100- to 1,000-fold). Similarly, TCA mutants exhibited AMP tolerance with a 100- to 1,000-fold increase in persister formation when challenged with LL-37 or human β-defensin 3 (hβD3). The ability of TCA cycle mutants to tolerate the innate immune system was further examined with a D. melanogaster model. Both males and females infected with TCA cycle mutants exhibited increased mortality and had higher bacterial burdens (1.5 log) during the course of the infection. These results suggest increasing the percentage of persister cells leads to increased tolerance to components of the innate immune system.

| [1] |
Lowy FD (1998) Staphylococcus aureus infections. N Engl J Med 339: 520-532. doi: 10.1056/NEJM199808203390806

|
| [2] |
Chang FY, Peacock JE, Musher DM, et al. (2003) Staphylococcus aureus bacteremia: recurrence and the impact of antibiotic treatment in a prospective multicenter study. Medicine (Baltimore) 82: 333-339. doi: 10.1097/01.md.0000091184.93122.09
|
| [3] |
Fowler VG, Boucher HW, Corey GR, et al. (2006) Daptomycin versus standard therapy for bacteremia and endocarditis caused by Staphylococcus aureus. N Engl J Med 355: 653-665. doi: 10.1056/NEJMoa053783
|
| [4] |
Welsh KJ, Skrobarcek KA, Abbott AN, et al. (2011) Predictors of relapse of methicillin-resistant Staphylococcus aureus bacteremia after treatment with vancomycin. J Clin Microbiol 49: 3669-3672. doi: 10.1128/JCM.05287-11
|
| [5] | Wolcott RD, Rhoads DD, Bennett ME, et al. (2010) Chronic wounds and the medical biofilm paradigm. J Wound Care 19: 45-6, 48-50, 52-3. |
| [6] |
Lewis K (2010) Persister cells. Annu Rev Microbiol 64: 357-372. doi: 10.1146/annurev.micro.112408.134306
|
| [7] |
Lewis K (2013) Platforms for antibiotic discovery. Nat Rev Drug Discov 12: 371-387. doi: 10.1038/nrd3975
|
| [8] |
Conlon BP, Rowe SE, Gandt AB, et al. (2016) Persister formation in Staphylococcus aureus is associated with ATP depletion. Nat Microbiol 1: 16051. doi: 10.1038/nmicrobiol.2016.51
|
| [9] |
Stapels DAC, Hill PWS, Westermann AJ, et al. (2018) Salmonella persisters undermine host immune defenses during antibiotic treatment. Science 362: 1156-1160. doi: 10.1126/science.aat7148
|
| [10] |
Thurlow LR, Hanke ML, Fritz T, et al. (2011) Staphylococcus aureus biofilms prevent macrophage phagocytosis and attenuate inflammation in vivo. J Immunol 186: 6585-6596. doi: 10.4049/jimmunol.1002794
|
| [11] |
Rowe SE, Wagner NJ, Li L, et al. (2020) Reactive oxygen species induce antibiotic tolerance during systemic Staphylococcus aureus infection. Nat Microbiol 5: 282-290. doi: 10.1038/s41564-019-0627-y
|
| [12] |
Peyrusson F, Varet H, Nguyen TK, et al. (2020) Intracellular Staphylococcus aureus persisters upon antibiotic exposure. Nat Commun 11: 2200. doi: 10.1038/s41467-020-15966-7
|
| [13] |
Zalis EA, Nuxoll AS, Manuse S, et al. (2019) Stochastic variation in expression of the tricarboxylic acid cycle produces persister Cells. mBio 10. doi: 10.1128/mBio.01930-19
|
| [14] |
Sieprawska-Lupa M, Mydel P, Krawczyk K, et al. (2004) Degradation of human antimicrobial peptide LL-37 by Staphylococcus aureus-derived proteinases. Antimicrob Agents Chemother 48: 4673-4679. doi: 10.1128/AAC.48.12.4673-4679.2004
|
| [15] |
Neville F, Cahuzac M, Konovalov O, et al. (2006) Lipid headgroup discrimination by antimicrobial peptide LL-37: insight into mechanism of action. Biophys J 90: 1275-1287. doi: 10.1529/biophysj.105.067595
|
| [16] |
Sass V, Schneider T, Wilmes M, et al. (2010) Human beta-defensin 3 inhibits cell wall biosynthesis in Staphylococci. Infect Immun 78: 2793-2800. doi: 10.1128/IAI.00688-09
|
| [17] |
Dushay MS, Eldon ED (1998) Drosophila immune responses as models for human immunity. Am J Hum Genet 62: 10-14. doi: 10.1086/301694
|
| [18] |
Salminen TS, Vale PF (2020) Drosophila as a model system to investigate the effects of mitochondrial variation on innate immunity. Front Immunol 11: 521. doi: 10.3389/fimmu.2020.00521
|
| [19] |
Marr AK, Gooderham WJ, Hancock RE (2006) Antibacterial peptides for therapeutic use: obstacles and realistic outlook. Curr Opin Pharmacol 6: 468-472. doi: 10.1016/j.coph.2006.04.006
|
| [20] |
Osei Sekyere J, Amoako DG (2017) Carbonyl Cyanide m-Chlorophenylhydrazine (CCCP) reverses resistance to colistin, but not to carbapenems and tigecycline in multidrug-resistant Enterobacteriaceae. Front Microbiol 8: 228. doi: 10.3389/fmicb.2017.00228
|
| [21] |
Short SM, Wolfner MF, Lazzaro BP (2012) Female Drosophila melanogaster suffer reduced defense against infection due to seminal fluid components. J Insect Physiol 58: 1192-1201. doi: 10.1016/j.jinsphys.2012.06.002
|
| [22] |
Alborn WE, Allen NE, Preston DA (1991) Daptomycin disrupts membrane potential in growing Staphylococcus aureus. Antimicrob Agents Chemother 35: 2282-2287. doi: 10.1128/AAC.35.11.2282
|
| [23] |
Patel D, Husain M, Vidaillac C, et al. (2011) Mechanisms of in-vitro-selected daptomycin-non-susceptibility in Staphylococcus aureus. Int J Antimicrob Agents 38: 442-446. doi: 10.1016/j.ijantimicag.2011.06.010
|
| [24] |
Needham AJ, Kibart M, Crossley H, et al. (2004) Drosophila melanogaster as a model host for Staphylococcus aureus infection. Microbiology (Reading) 150: 2347-2355. doi: 10.1099/mic.0.27116-0
|
| [25] |
Garver LS, Wu J, Wu LP (2006) The peptidoglycan recognition protein PGRP-SC1a is essential for Toll signaling and phagocytosis of Staphylococcus aureus in Drosophila. Proc Natl Acad Sci USA 103: 660-665. doi: 10.1073/pnas.0506182103
|
| [26] |
Conlon BP, Nakayasu ES, Fleck LE, et al. (2013) Activated ClpP kills persisters and eradicates a chronic biofilm infection. Nature 503: 365-370. doi: 10.1038/nature12790
|
| [27] |
Kreiswirth BN, Lofdahl S, Betley MJ, et al. (1983) The toxic shock syndrome exotoxin structural gene is not detectably transmitted by a prophage. Nature 305: 709-712. doi: 10.1038/305709a0
|
| [28] |
Bottger EC, Springer B, Prammananan T, et al. (2001) Structural basis for selectivity and toxicity of ribosomal antibiotics. EMBO Rep 2: 318-323. doi: 10.1093/embo-reports/kve062
|
| [29] |
Hooper DC, Wolfson JS, McHugh GL, et al. (1982) Effects of novobiocin, coumermycin A1, clorobiocin, and their analogs on Escherichia coli DNA gyrase and bacterial growth. Antimicrob Agents Chemother 22: 662-671. doi: 10.1128/AAC.22.4.662
|
| [30] |
Shan Y, Brown Gandt A, Rowe SE, et al. (2017) ATP-dependent persister formation in Escherichia coli. mBio 8. doi: 10.1128/mBio.02267-16
|
| [31] |
Hancock RE (1997) Peptide antibiotics. Lancet 349: 418-422. doi: 10.1016/S0140-6736(97)80051-7
|
| [32] |
Wang Y, Bojer MS, George SE, et al. (2018) Inactivation of TCA cycle enhances Staphylococcus aureus persister cell formation in stationary phase. Sci Rep 8: 10849. doi: 10.1038/s41598-018-29123-0
|
| [33] |
Zheng EJ, Stokes JM, Collins JJ (2020) Eradicating bacterial persisters with combinations of strongly and weakly metabolism-dependent antibiotics. Cell Chem Biol 27: 1544-1552 e1543. doi: 10.1016/j.chembiol.2020.08.015
|
| [34] |
Midorikawa K, Ouhara K, Komatsuzawa H, et al. (2003) Staphylococcus aureus susceptibility to innate antimicrobial peptides, beta-defensins and CAP18, expressed by human keratinocytes. Infect Immun 71: 3730-3739. doi: 10.1128/IAI.71.7.3730-3739.2003
|
| [35] |
McKean KA, Nunney L (2001) Increased sexual activity reduces male immune function in Drosophila melanogaster. Proc Natl Acad Sci USA 98: 7904-7909. doi: 10.1073/pnas.131216398
|
| [36] |
Garcia-Lara J, Needham AJ, Foster SJ (2005) Invertebrates as animal models for Staphylococcus aureus pathogenesis: a window into host-pathogen interaction. FEMS Immunol Med Microbiol 43: 311-323. doi: 10.1016/j.femsim.2004.11.003
|
| [37] |
McGraw LA, Gibson G, Clark AG, et al. (2004) Genes regulated by mating, sperm, or seminal proteins in mated female Drosophila melanogaster. Curr Biol 14: 1509-1514. doi: 10.1016/j.cub.2004.08.028
|
| [38] |
Domanitskaya EV, Liu H, Chen S, et al. (2007) The hydroxyproline motif of male sex peptide elicits the innate immune response in Drosophila females. FEBS J 274: 5659-5668. doi: 10.1111/j.1742-4658.2007.06088.x
|
| [39] |
Wigby S, Domanitskaya EV, Choffat Y, et al. (2008) The effect of mating on immunity can be masked by experimental piercing in female Drosophila melanogaster. J Insect Physiol 54: 414-420. doi: 10.1016/j.jinsphys.2007.10.010
|
| [40] |
Pitnick S, Markow TA, Spicer GS (1995) Delayed male maturity is a cost of producing large sperm in Drosophila. Proc Natl Acad Sci U S A 92: 10614-10618. doi: 10.1073/pnas.92.23.10614
|
| [41] |
McRobert SP, Tompkins L (1988) Two consequences of homosexual courtship performed by Drosophila Melanogaster and Drosophila Affinis males. Evolution 42: 1093-1097. doi: 10.1111/j.1558-5646.1988.tb02528.x
|
| [42] |
Lehrer RI, Ganz T (1999) Antimicrobial peptides in mammalian and insect host defence. Curr Opin Immunol 11: 23-27. doi: 10.1016/S0952-7915(99)80005-3
|
| [43] |
Kocks C, Cho JH, Nehme N, et al. (2005) Eater, a transmembrane protein mediating phagocytosis of bacterial pathogens in Drosophila. Cell 123: 335-346. doi: 10.1016/j.cell.2005.08.034
|
 microbiol-07-04-031-s001.pdf microbiol-07-04-031-s001.pdf |
 |





Figures(6)

Alexis M. Hobbs, Kennedy E. Kluthe, Kimberly A. Carlson, Austin S. Nuxoll. Interruption of the tricarboxylic acid cycle in Staphylococcus aureus leads to increased tolerance to innate immunity[J]. AIMS Microbiology, 2021, 7(4): 513-527. doi: 10.3934/microbiol.2021031







 DownLoad:
DownLoad: