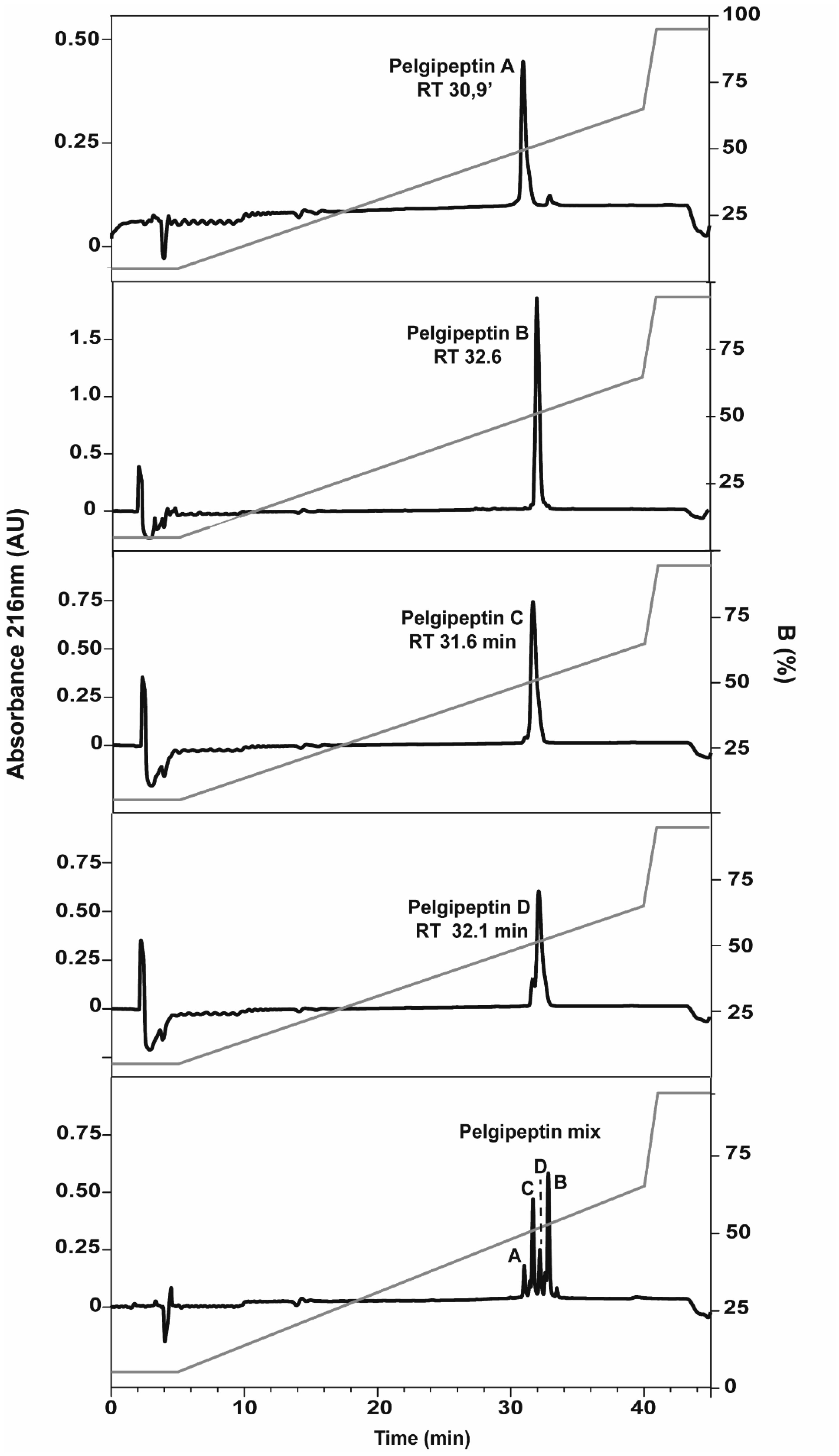
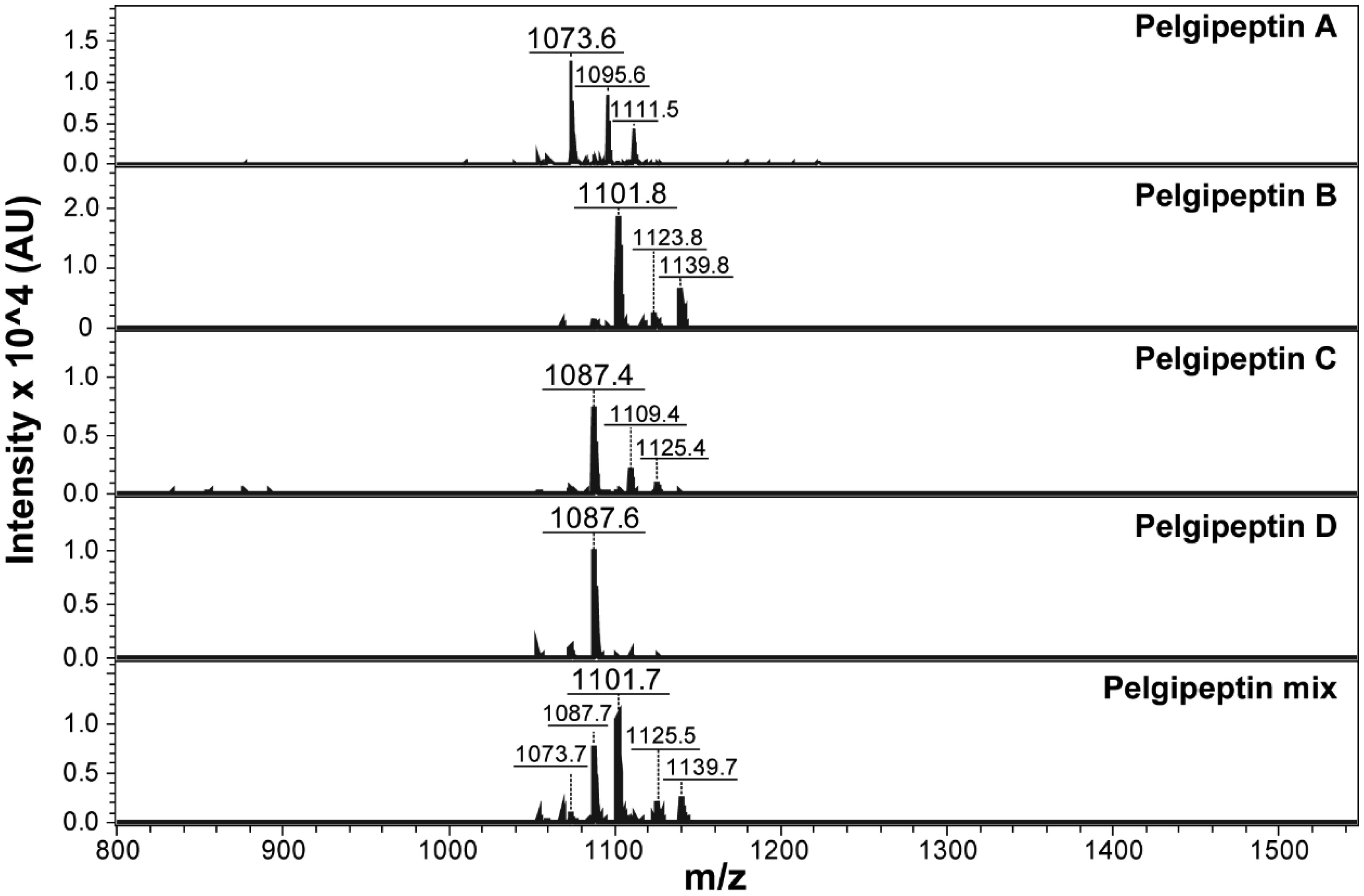
Systemic mycoses have become a major cause of morbidity and mortality, particularly among immunocompromised hosts and long-term hospitalized patients. Conventional antifungal agents are limited because of not only their costs and toxicity but also the rise of resistant strains. Lipopeptides from Paenibacillus species exhibit antimicrobial activity against a wide range of human and plant bacterial pathogens. However, the antifungal potential of these compounds against important human pathogens has not yet been fully evaluated, except for Candida albicans. Paenibacillus elgii produces a family of lipopeptides named pelgipeptins, which are synthesized by a non-ribosomal pathway, such as polymyxin. The present study aimed to evaluate the activity of pelgipeptins produced by P. elgii AC13 against Cryptococcus neoformans, Paracoccidioides brasiliensis, and Candida spp. Pelgipeptins were purified from P. elgii AC13 cultures and characterized by high-performance liquid chromatography (HPLC) and mass spectrometry (MALDI-TOF MS). The in vitro antifugal activity of pelgipeptins was evaluated against C. neoformans H99, P. brasiliensis PB18, C. albicans SC 5314, Candida glabrata ATCC 90030, and C. albicans biofilms. Furthermore, the minimal inhibitory concentration (MIC) was determined according to the CLSI microdilution method. Fluconazole and amphotericin B were also used as a positive control. Pelgipeptins A to D inhibited the formation and development of C. albicans biofilms and presented activity against all tested microorganisms. The minimum inhibitory concentration values ranged from 4 to 64 µg/mL, which are in the same range as fluconazole MICs. These results highlight the potential of pelgipeptins not only as antimicrobials against pathogenic fungi that cause systemic mycoses but also as coating agents to prevent biofilm formation on medical devices.
Citation: Débora Luíza Albano Fulgêncio, Rosiane Andrade da Costa, Fernanda Guilhelmelli, Calliandra Maria de Souza Silva, Daniel Barros Ortega, Thiago Fellipe de Araujo, Philippe Spezia Silva, Ildinete Silva-Pereira, Patrícia Albuquerque, Cristine Chaves Barreto. In vitro antifungal activity of pelgipeptins against human pathogenic fungi and Candida albicans biofilms[J]. AIMS Microbiology, 2021, 7(1): 28-39. doi: 10.3934/microbiol.2021003
Systemic mycoses have become a major cause of morbidity and mortality, particularly among immunocompromised hosts and long-term hospitalized patients. Conventional antifungal agents are limited because of not only their costs and toxicity but also the rise of resistant strains. Lipopeptides from Paenibacillus species exhibit antimicrobial activity against a wide range of human and plant bacterial pathogens. However, the antifungal potential of these compounds against important human pathogens has not yet been fully evaluated, except for Candida albicans. Paenibacillus elgii produces a family of lipopeptides named pelgipeptins, which are synthesized by a non-ribosomal pathway, such as polymyxin. The present study aimed to evaluate the activity of pelgipeptins produced by P. elgii AC13 against Cryptococcus neoformans, Paracoccidioides brasiliensis, and Candida spp. Pelgipeptins were purified from P. elgii AC13 cultures and characterized by high-performance liquid chromatography (HPLC) and mass spectrometry (MALDI-TOF MS). The in vitro antifugal activity of pelgipeptins was evaluated against C. neoformans H99, P. brasiliensis PB18, C. albicans SC 5314, Candida glabrata ATCC 90030, and C. albicans biofilms. Furthermore, the minimal inhibitory concentration (MIC) was determined according to the CLSI microdilution method. Fluconazole and amphotericin B were also used as a positive control. Pelgipeptins A to D inhibited the formation and development of C. albicans biofilms and presented activity against all tested microorganisms. The minimum inhibitory concentration values ranged from 4 to 64 µg/mL, which are in the same range as fluconazole MICs. These results highlight the potential of pelgipeptins not only as antimicrobials against pathogenic fungi that cause systemic mycoses but also as coating agents to prevent biofilm formation on medical devices.

| [1] |
Calderone RA, Fonzi WA (2001) Virulence factors of Candida albicans. Trends Microbiol 9: 327-335. doi: 10.1016/S0966-842X(01)02094-7

|
| [2] |
Shikanai-Yasuda MA (2015) Paracoccidioidomycosis treatment. Rev Inst Med Trop Sao Paulo 57: 31-37. doi: 10.1590/S0036-46652015000700007
|
| [3] |
Bongomin F, Gago S, Oladele RO, et al. (2017) Global and multi-national prevalence of fungal diseases—estimate precision. J Fungi 3: 57. doi: 10.3390/jof3040057
|
| [4] |
Sudbery PE (2011) Growth of Candida albicans hyphae. Nat Rev Microbiol 9: 737-748. doi: 10.1038/nrmicro2636
|
| [5] |
Horn DL, Neofytos D, Anaissie EJ, et al. (2009) Epidemiology and outcomes of candidemia in 2019 patients: data from the prospective antifungal therapy alliance registry. Clin Infect Dis 48: 1695-1703. doi: 10.1086/599039
|
| [6] |
Ramage G, Mowat E, Jones B, et al. (2009) Our current understanding of fungal biofilms. Crit Rev Microbiol 35: 340-355. doi: 10.3109/10408410903241436
|
| [7] |
Levitz SM (1991) The ecology of Cryptococcus neoformans and the epidemiology of cryptococcosis. Rev Infect Dis 13: 1163-1169. doi: 10.1093/clinids/13.6.1163
|
| [8] |
Idnurm A, Bahn YS, Nielsen K, et al. (2005) Deciphering the model pathogenic fungus Cryptococcus neoformans. Nat Rev Microbiol 3: 753-764. doi: 10.1038/nrmicro1245
|
| [9] |
Velagapudi R, Hsueh YP, Geunes-Boyer S, et al. (2009) Spores as infectious propagules of Cryptococcus neoformans. Infect Immun 77: 4345-4355. doi: 10.1128/IAI.00542-09
|
| [10] |
Rajasingham R, Smith RM, Park BJ, et al. (2017) Global burden of disease of HIV-associated cryptococcal meningitis: an updated analysis. Lancet Infect Dis 17: 873-881. doi: 10.1016/S1473-3099(17)30243-8
|
| [11] |
San-Blas F, Cova LJ (1975) Growth curves of the yeast-like form of Paracocidioides brasiliensis. Sabouraudia 1: 22-29. doi: 10.1080/00362177585190041
|
| [12] |
San-Blas G (1985) Paracoccidioides brasiliensis: Cell Wall Glucans, Pathogenicity, and Dimorphism. Current Topics in Medical Mycology New York: Springer, 235-257. doi: 10.1007/978-1-4613-9547-8_9
|
| [13] |
Mendes RP, Cavalcante R de S, Marques SA, et al. (2017) Paracoccidioidomycosis: Current perspectives from Brazil. Open Microbiol J 11: 224-282. doi: 10.2174/1874285801711010224
|
| [14] | da Costa MM, Marques da Silva SH (2014) Epidemiology, clinical, and therapeutic aspects of Paracoccidioidomycosis. Curr Trop Med Reports 1: 138-144. |
| [15] |
Stein T (2005) Bacillus subtilis antibiotics: structures, syntheses and specific functions. Mol Microbiol 56: 845-857. doi: 10.1111/j.1365-2958.2005.04587.x
|
| [16] |
Cochrane SA, Vederas JC (2016) Lipopeptides from Bacillus and Paenibacillus spp.: a gold mine of antibiotic candidates. Med Res Rev 36: 4-31. doi: 10.1002/med.21321
|
| [17] |
Meena KR, Kanwar SS (2015) Lipopeptides as the antifungal and antibacterial agents: applications in food safety and therapeutics. Biomed Res Int 2015: 473050. doi: 10.1155/2015/473050
|
| [18] |
Olishevska S, Nickzad A, Déziel E (2019) Bacillus and Paenibacillus secreted polyketides and peptides involved in controlling human and plant pathogens. Appl Microbiol Biotechnol 103: 1189-1215. doi: 10.1007/s00253-018-9541-0
|
| [19] |
Velkov T, Roberts KD, Nation RL, et al. (2013) Pharmacology of polymyxins: new insights into an ‘old’ class of antibiotics. Future Microbiol 8: 711-724. doi: 10.2217/fmb.13.39
|
| [20] |
Wu XC, Shen XB, Ding R, et al. (2010) Isolation and partial characterization of antibiotics produced by Paenibacillus elgii B69. FEMS Microbiol Lett 310: 32-38. doi: 10.1111/j.1574-6968.2010.02040.x
|
| [21] |
Ding R, Wu XC, Qian CD, et al. (2011) Isolation and identification of lipopeptide antibiotics from Paenibacillus elgii B69 with inhibitory activity against methicillin-resistant Staphylococcus aureus. J Microbiol 49: 942-949. doi: 10.1007/s12275-011-1153-7
|
| [22] |
Costa R, Ortega D, Fulgêncio D, et al. (2019) Checkerboard testing method indicates synergic effect of pelgipeptins against multidrug resistant Klebsiella pneumoniae. Biotechnol Res Innov 3: 187-191. doi: 10.1016/j.biori.2018.12.001
|
| [23] |
Ortega DB, Costa RA, Pires AS, et al. (2018) Draft Genome sequence of the antimicrobial-producing strain Paenibacillus elgii AC13. Genome Announc 6: e00573-18. doi: 10.1128/genomeA.00573-18
|
| [24] |
Murphy JB, Kies MW (1960) Note on spectrophotometric determination of proteins in dilute solutions. Biochim Biophys Acta 45: 382-384. doi: 10.1016/0006-3002(60)91464-5
|
| [25] | (2017) CLSIReference method for broth dilution antifungal susceptibility testing of yeasts CLSI Standard M27. Wayne, PA. |
| [26] | De-Souza-Silva CM, Guilhelmelli F, Zamith-Miranda D, et al. (2018) Broth microdilution in vitro screening: An easy and fast method to detect new antifungal compounds. J Vis Exp 132: 57127. |
| [27] |
Pankey G, Ashcraft D, Kahn H, et al. (2014) Time-kill assay and etest evaluation for synergy with polymyxin B and fluconazole against Candida glabrata. Antimicrob Agents Chemother 58: 5795-5800. doi: 10.1128/AAC.03035-14
|
| [28] |
Hsu LH, Wang HF, Sun PL, et al. (2017) The antibiotic polymyxin B exhibits novel antifungal activity against Fusarium species. Int J Antimicrob Agents 49: 740-748. doi: 10.1016/j.ijantimicag.2017.01.029
|
| [29] |
De Paula E Silva ACA, Oliveira HC, Silva JF, et al. (2013) Microplate alamarblue assay for paracoccidioides susceptibility testing. J Clin Microbiol 51: 1250-1252. doi: 10.1128/JCM.02914-12
|
| [30] |
Pierce CG, Uppuluri P, Tristan AR, et al. (2008) A simple and reproducible 96-well plate-based method for the formation of fungal biofilms and its application to antifungal susceptibility testing. Nat Protoc 3: 1494-1500. doi: 10.1038/nprot.2008.141
|
| [31] | Pirri G, Giuliani A, Nicoletto SF, et al. (2009) Lipopeptides as anti-infectives: a practical perspective. Cent Eur J Biol 4: 258-273. |
| [32] |
Trimble MJ, Mlynárčik P, Kolář M, et al. (2016) Polymyxin: Alternative mechanisms of action and resistance. Cold Spring Harb Perspect Med 6: 1-23. doi: 10.1101/cshperspect.a025288
|
| [33] |
Zhai B, Zhou H, Yang L, et al. (2010) Polymyxin B, in combination with fluconazole, exerts a potent fungicidal effect. J Antimicrob Chemother 65: 931-938. doi: 10.1093/jac/dkq046
|
| [34] |
Qian CD, Liu TZ, Zhou SL, et al. (2012) Identification and functional analysis of gene cluster involvement in biosynthesis of the cyclic lipopeptide antibiotic pelgipeptin produced by Paenibacillus elgii. BMC Microbiol 12: 197. doi: 10.1186/1471-2180-12-197
|
| [35] |
Campoy S, Adrio JL (2017) Antifungals. Biochem Pharmacol 133: 86-96. doi: 10.1016/j.bcp.2016.11.019
|
| [36] |
Kim J, Il Kim P, Bong KM, et al. (2018) Isolation and structural elucidation of pelgipeptin E, a novel pore-forming pelgipeptin analog from Paenibacillus elgii with low hemolytic activity. J Antibiot (Tokyo) 71: 1008-1017. doi: 10.1038/s41429-018-0095-2
|
| [37] |
Bachmann SP, VandeWalle K, Ramage G, et al. (2002) In vitro activity of caspofungin against Candida albicans biofilms. Antimicrob Agents Chemother 46: 3591-3596. doi: 10.1128/AAC.46.11.3591-3596.2002
|
| [38] |
Bruzual I, Riggle P, Hadley S, et al. (2007) Biofilm formation by fluconazole-resistant Candida albicans strains is inhibited by fluconazole. J Antimicrob Chemother 59: 441-450. doi: 10.1093/jac/dkl521
|
| [39] |
Ramage G, Vande Walle K, Wickes BL, et al. (2001) Standardized method for in vitro antifungal susceptibility testing of Candida albicans biofilms. Antimicrob Agents Chemother 45: 2475-2479. doi: 10.1128/AAC.45.9.2475-2479.2001
|
| [40] |
Ramage G, VandeWalle K, Lopez-Ribot J, et al. (2002) The filamentation pathway controlled by the Efg1 regulator protein is required for normal biofilm formation and development in Candida albicans. FEMS Microbiol Lett 214: 95-100. doi: 10.1111/j.1574-6968.2002.tb11330.x
|
Figures(2) / Tables(3)

Débora Luíza Albano Fulgêncio, Rosiane Andrade da Costa, Fernanda Guilhelmelli, Calliandra Maria de Souza Silva, Daniel Barros Ortega, Thiago Fellipe de Araujo, Philippe Spezia Silva, Ildinete Silva-Pereira, Patrícia Albuquerque, Cristine Chaves Barreto. In vitro antifungal activity of pelgipeptins against human pathogenic fungi and Candida albicans biofilms[J]. AIMS Microbiology, 2021, 7(1): 28-39. doi: 10.3934/microbiol.2021003







 DownLoad:
DownLoad: