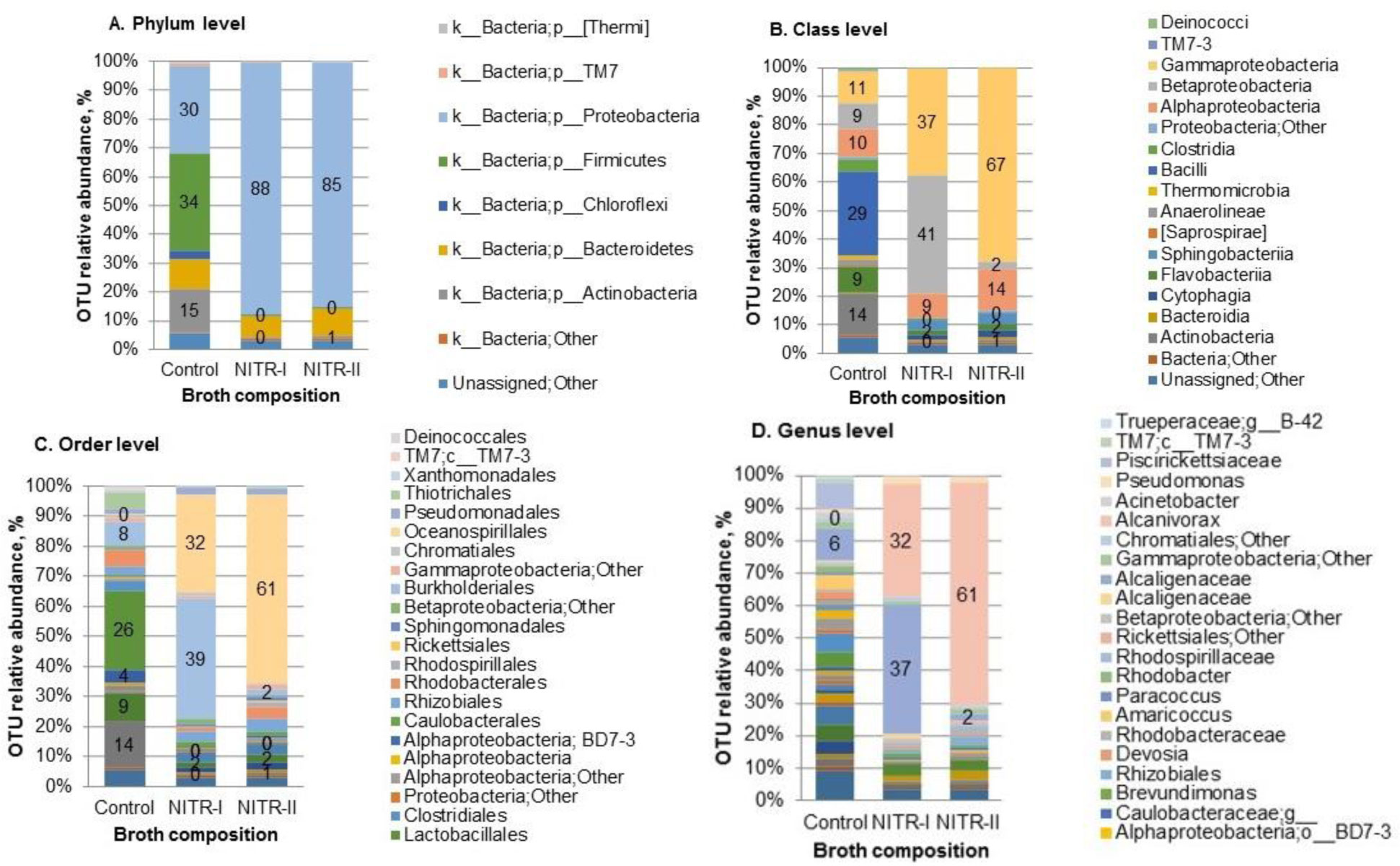
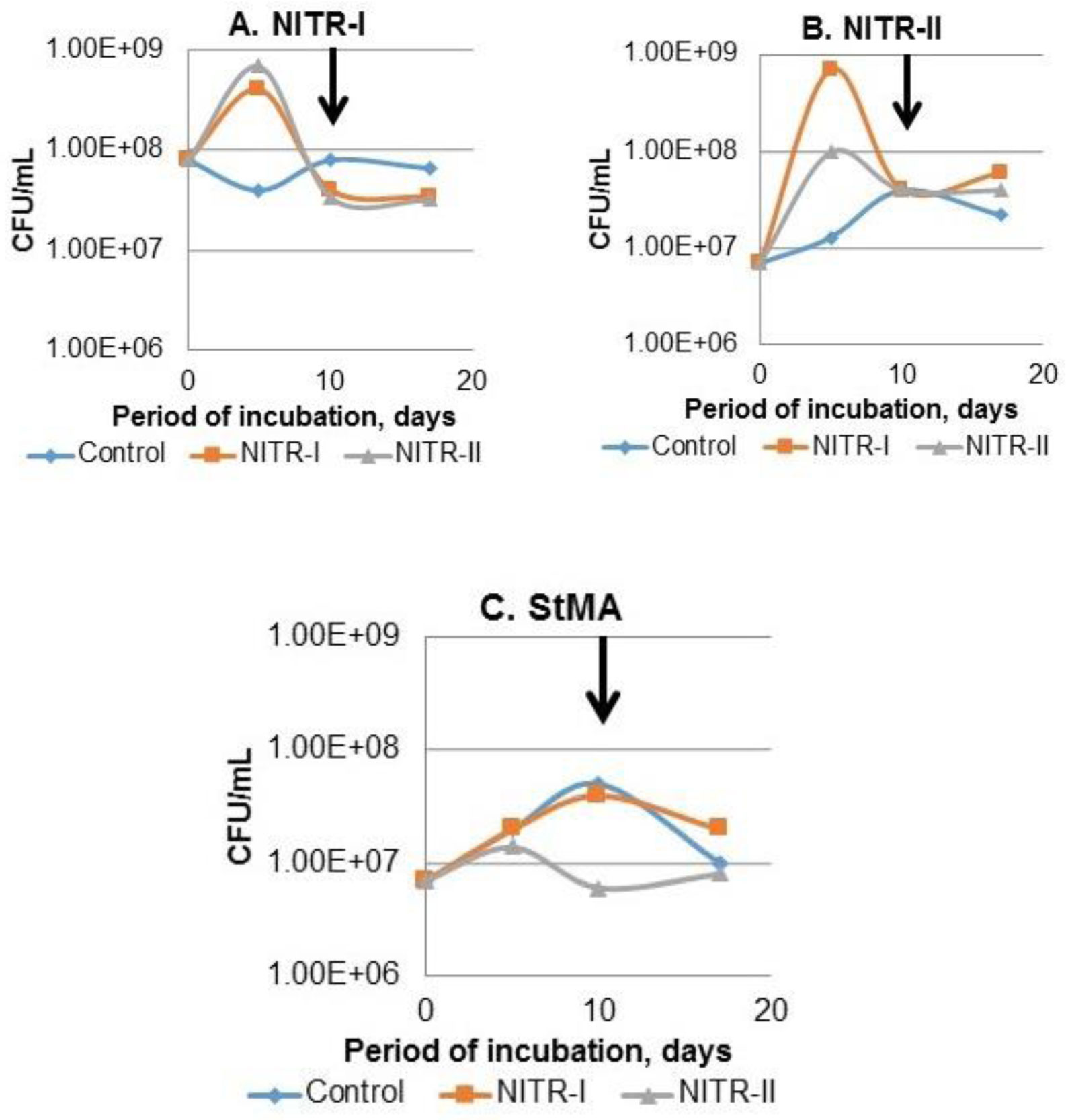
Removal of nitrogen from wastewaters (WW) represents a global problem. The low nitrification rate during WW treatment is often caused by ecotoxicity. This problem is attributed mostly to the industrial WW. Our study was focused on the testing of industrial WW and activated sludge (AS) with the aim to reveal the abundance of nitrifiers and increase their biomass, thus, providing the additional step, i.e., bioaugmentation, within the technological process of WW treatment. Plating of AS on the selective solidified media designated for the 1st and 2nd nitrification stages, resulted in the shift in bacterial community structure with dominated Alcaligenaceae and Alcanivorax for the 1st stage, and Alcanivorax-for the 2nd stage of nitrification, respectively. Incubation of AS in the presence of real WW and selective nitrification broth resulted in a considerable increase (one or two magnitudes in the presence of the 1st and 2nd stage nitrification broth, respectively) of culturable nitrifiers after 5 days incubation under aerated conditions. The obtained data provide with evidence about a possibility to strengthen the role of heterotrophic nitrifiers in the treatment of industrial WW, where toxicity obstacles inhibited nitrification under conventional conditions.
Citation: Mārtiņš Kalniņš, Andrejs Bērziņš, Dita Gudrā, Kaspars Megnis, Dāvids Fridmanis, Pavel Danilko, Olga Muter. Selective enrichment of heterotrophic nitrifiers Alcaligenaceae and Alcanivorax spp. from industrial wastewaters[J]. AIMS Microbiology, 2020, 6(1): 32-42. doi: 10.3934/microbiol.2020002
Removal of nitrogen from wastewaters (WW) represents a global problem. The low nitrification rate during WW treatment is often caused by ecotoxicity. This problem is attributed mostly to the industrial WW. Our study was focused on the testing of industrial WW and activated sludge (AS) with the aim to reveal the abundance of nitrifiers and increase their biomass, thus, providing the additional step, i.e., bioaugmentation, within the technological process of WW treatment. Plating of AS on the selective solidified media designated for the 1st and 2nd nitrification stages, resulted in the shift in bacterial community structure with dominated Alcaligenaceae and Alcanivorax for the 1st stage, and Alcanivorax-for the 2nd stage of nitrification, respectively. Incubation of AS in the presence of real WW and selective nitrification broth resulted in a considerable increase (one or two magnitudes in the presence of the 1st and 2nd stage nitrification broth, respectively) of culturable nitrifiers after 5 days incubation under aerated conditions. The obtained data provide with evidence about a possibility to strengthen the role of heterotrophic nitrifiers in the treatment of industrial WW, where toxicity obstacles inhibited nitrification under conventional conditions.

| [1] |
Sahrawat KL (2008) Factors affecting nitrification in soils. Commun Soil Sci Plan 39: 1436-1446. doi: 10.1080/00103620802004235

|
| [2] | Stein LY (2014) Heterotrophic nitrification and nitrifier denitrification. Nitrification . |
| [3] | Thymann A (2013) The co-existence of heterotrophs and nitrifiers in nitrifying biofilm a study on the relationship between heterotrophic and nitrifying bacteria in a marine environment. Available from: https://rucforsk.ruc.dk/ws/portalfiles/portal/57744106/THE_CO-EXISTENCE_OF_HETEROTROPHS_AND_NITRIFIERS_IN_NITRIFYING_BIOFILM.pdf. |
| [4] |
Onyla CO, Uyub AM, Akunna JC, et al. (2001) Increasing the fertilizer value of palm oil mill sludge: Bioaugmentation in nitrification. Water Sci Technol 44: 157-162. doi: 10.2166/wst.2001.0608
|
| [5] |
Elmerich C (2002) Nitrification and denitrification in the activated sludge process. Res Microbiol 153: 253. doi: 10.1016/S0923-2508(02)01315-3
|
| [6] |
Velusamy K, Krishnani KK (2013) Heterotrophic nitrifying and oxygen tolerant denitrifying bacteria from greenwater system of coastal aquaculture. Appl Biochem Biotechnol 169: 1978-1992. doi: 10.1007/s12010-013-0109-2
|
| [7] |
Ge S, Wang S, Yang X, et al. (2015) Detection of nitrifiers and evaluation of partial nitrification for wastewater treatment: a review. Chemosphere 140: 85-98. doi: 10.1016/j.chemosphere.2015.02.004
|
| [8] |
Booth MS, Stark JM, Rastetter E (2005) Controls on nitrogen cycling in terrestrial ecosystems: A synthetic analysis of literature data. Ecol Monogr 75: 139-157. doi: 10.1890/04-0988
|
| [9] |
Iannacone F, Di Capua F, Granata F, et al. (2019) Effect of carbon-to-nitrogen ratio on simultaneous nitrification denitrification and phosphorus removal in a microaerobic moving bed biofilm reactor. J Environ Manage 250: 109518. doi: 10.1016/j.jenvman.2019.109518
|
| [10] |
Hu Z, Chandran K, Grasso D, et al. (2004) Comparison of nitrification inhibition by metals in batch and continuous flow reactors. Water Res 38: 3949-3959. doi: 10.1016/j.watres.2004.06.025
|
| [11] |
You SJ, Tsai YP, Huang RY (2009) Effect of heavy metals on nitrification performance in different activated sludge processes. J Hazard Mater 165: 987-994. doi: 10.1016/j.jhazmat.2008.10.112
|
| [12] |
Papirio S, Zou G, Ylinen A, et al. (2014) Effect of arsenic on nitrification of simulated mining water. Bioresour Technol 164: 149-154. doi: 10.1016/j.biortech.2014.04.072
|
| [13] |
di Biase A, et al. (2020) Ammonia, thiocyanate, and cyanate removal in an aerobic up-flow submerged attached growth reactor treating gold mine wastewater. Chemosphere 243: 125395. doi: 10.1016/j.chemosphere.2019.125395
|
| [14] |
Tang HL, Chen H (2015) Nitrification at full-scale municipal wastewater treatment plants: Evaluation of inhibition and bioaugmentation of nitrifiers. Bioresour Technol 190: 76-81. doi: 10.1016/j.biortech.2015.04.063
|
| [15] |
Parker D, Wanner J (2007) Review of methods for improving nitrification through bioaugmentation. Water Pract 1: 1-16. doi: 10.2175/193317707X256964
|
| [16] | Zimmerman RA, Bradshaw AT, Richard D (2004) Acclimation of nitrifiers for activated sludge treatment: a bench scale evaluation. Water Intell Online 3. |
| [17] |
Hayatsu M, Tago K, Saito M (2008) Various players in the nitrogen cycle: Diversity and functions of the microorganisms involved in nitrification and denitrification. Soil Sci Plant Nutr 54: 33-45. doi: 10.1111/j.1747-0765.2007.00195.x
|
| [18] |
Bartrolí A, Carrera J, Pérez J (2011) Bioaugmentation as a tool for improving the start-up and stability of a pilot-scale partial nitrification biofilm airlift reactor. Bioresour Technol 102: 4370-4375. doi: 10.1016/j.biortech.2010.12.084
|
| [19] |
Mannucci A, Munz G, Mori G, et al. (2015) Modeling bioaugmentation with nitrifiers in membrane bioreactors. Water Sci Technol 71: 15-21. doi: 10.2166/wst.2014.456
|
| [20] |
Figdore BA, Stensel HD, Winkler MKH, et al. (2018) Comparison of different aerobic granular sludge types for activated sludge nitrification bioaugmentation potential. Bioresour Technol 251: 189-196. doi: 10.1016/j.biortech.2017.11.004
|
| [21] |
Li AJ, Li XY, Yu HQ (2013) Aerobic sludge granulation facilitated by activated carbon for partial nitrification treatment of ammonia-rich wastewater. Chem Eng J 218: 253-259. doi: 10.1016/j.cej.2012.12.044
|
| [22] | Yu L, Peng D, Pan R (2012) Shifts in nitrification kinetics and microbial community during bioaugmentation of activated sludge with nitrifiers enriched on sludge reject water. J Biomed Biotechnol 2012: 691894. |
| [23] |
Pfeltz RF, Schmidt JL, Wilkinson BJ (2001) A microdilution plating method for population analysis of antibiotic-resistant staphylococci. Microb Drug Resist 7: 289-295. doi: 10.1089/10766290152652846
|
| [24] | Kalnins MO (2019) Application of enzyme activity assay for an improved enumeration of culturable nitrifiers. Environ Exp Biol 17: 49. |
| [25] |
Camiña F, Trasar-Cepeda C, Gil-Sotres F, et al. (1998) Measurement of dehydrogenase activity in acid soils rich in organic matter. Soil Biol Biochem 30: 1005-1011. doi: 10.1016/S0038-0717(98)00010-8
|
| [26] |
Milani C, Hevia A, Foroni E, et al. (2013) Assessing the fecal mcrobiota: an optimized ion torrent 16S rRNA gene-based analysis protocol. PLoS One 8: e68739. doi: 10.1371/journal.pone.0068739
|
| [27] |
Pylro VS, Roesch LFW, Morais DK, et al. (2014) Data analysis for 16S microbial profiling from different benchtop sequencing platforms. J Microbiol Methods 107: 30-37. doi: 10.1016/j.mimet.2014.08.018
|
| [28] |
Edgar RC (2010) Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26: 2460-2461. doi: 10.1093/bioinformatics/btq461
|
| [29] |
DeSantis TZ, Hugenholtz P, Larsen N, et al. (2006) Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol 72: 5069-5072. doi: 10.1128/AEM.03006-05
|
| [30] |
Yu K, Zhang T (2012) Metagenomic and metatranscriptomic analysis of microbial community structure and gene expression of activated sludge. PLoS One 7: e38183. doi: 10.1371/journal.pone.0038183
|
| [31] |
Wang X, Wen X, Criddle C, et al. (2010) Community analysis of ammonia-oxidizing bacteria in activated sludge of eight wastewater treatment systems. J Environ Sci 22: 627-634. doi: 10.1016/S1001-0742(09)60155-8
|
| [32] |
Grunditz C, Dalhammar G (2001) Development of nitrification inhibition assays using pure cultures of Nitrosomonas and Nitrobacter. Water Res 35: 433-440. doi: 10.1016/S0043-1354(00)00312-2
|
| [33] |
Ye L, Shao MF, Zhang T, et al. (2011) Analysis of the bacterial community in a laboratory-scale nitrification reactor and a wastewater treatment plant by 454-pyrosequencing. Water Res 45: 4390-4398. doi: 10.1016/j.watres.2011.05.028
|
| [34] |
Pollock J, Glendinning L, Wisedchanwet T, et al. (2018) The madness of microbiome: Attempting to find consensus ‘best practice’ for 16S microbiome studies. Appl Environ Microbiol 84: e02627-17. doi: 10.1128/AEM.02627-17
|
| [35] | Hornung BVH, Zwittink RD, Kuijper EJ (2019) Issues and current standards of controls in microbiome research. FEMS Microbiol Ecol 95: fiz045. |
| [36] |
Satoh H, Nakamura Y, Ono H, et al. (2003) Effect of oxygen concentration on nitrification and denitrification in single activated sludge flocs. Biotechnol Bioeng 83: 604-607. doi: 10.1002/bit.10717
|
| [37] |
Hocaoglu SM, Insel G, Cokgor UU, et al. (2011) Effect of low dissolved oxygen on simultaneous nitrification and denitrification in a membrane bioreactor treating black water. Bioresour Technol 102: 4333-4340. doi: 10.1016/j.biortech.2010.11.096
|
| [38] | Chen Q, Ni J (2012) Ammonium removal by Agrobacterium sp. LAD9 capable of heterotrophic nitrification-aerobic denitrification. J Biosci Bioeng 102: 4333-4340. |
| [39] |
Ren YX, Yang L, Liang X (2014) The characteristics of a novel heterotrophic nitrifying and aerobic denitrifying bacterium, Acinetobacter junii YB. Bioresour Technol 171: 1-9. doi: 10.1016/j.biortech.2014.08.058
|
| [40] |
Zhao B, He YL, Hughes J, et al. (2010) Heterotrophic nitrogen removal by a newly isolated Acinetobacter calcoaceticus HNR. Bioresour Technol 101: 5194-5200. doi: 10.1016/j.biortech.2010.02.043
|
| [41] |
Sotres A, Cerrillo M, Viñas M, et al. (2016) Nitrogen removal in a two-chambered microbial fuel cell: Establishment of a nitrifying-denitrifying microbial community on an intermittent aerated cathode. Chem Eng J 284: 905-916. doi: 10.1016/j.cej.2015.08.100
|
| [42] | Shoda M (2017) Heterotrophic nitrification and aerobic denitrification by Alcaligenes faecalis No. 4. Nitrification and Denitrification . |
| [43] |
Honda N, Hirai M, Ano T, et al. (1999) Control of tomato damping-off caused by Rhizoctonia solani by the heterotrophic nitrifier Alcaligenes faecalis and its product, hydroxylamine. Japanese J Phytopathol 65: 153-162. doi: 10.3186/jjphytopath.65.153
|
| [44] |
Liu C, Shao Z (2005) Alcanivorax dieselolei sp. nov., a novel alkane-degrading bacterium isolated from sea water and deep-sea sediment. Int J Syst Evol Microbiol 55: 1181-1186. doi: 10.1099/ijs.0.63443-0
|
| [45] |
Li H, Wang X, Mu B, et al. (2013) Molecular detection, quantification and distribution of alkane-degrading bacteria in production water from low temperature oilfields. Int Biodeterior Biodegrad 76: 49-57. doi: 10.1016/j.ibiod.2012.06.007
|
| [46] |
Nakano M, Shimizu Y, Okumura H, et al. (2008) Construction of a consortium comprising ammonia-oxidizing bacteria and denitrifying bacteria isolated from marine sediment. Biocontrol Sci 13: 73-89. doi: 10.4265/bio.13.73
|
 microbiol-06-01-002-s001.pdf microbiol-06-01-002-s001.pdf |
 |
Figures(2) / Tables(1)

Mārtiņš Kalniņš, Andrejs Bērziņš, Dita Gudrā, Kaspars Megnis, Dāvids Fridmanis, Pavel Danilko, Olga Muter. Selective enrichment of heterotrophic nitrifiers Alcaligenaceae and Alcanivorax spp. from industrial wastewaters[J]. AIMS Microbiology, 2020, 6(1): 32-42. doi: 10.3934/microbiol.2020002







 DownLoad:
DownLoad: