Citation: Manuel Molina, Manuel Mota, Alfonso Ramos. Mathematical modeling in semelparous biological species through two-sex branching processes[J]. Mathematical Biosciences and Engineering, 2024, 21(6): 6407-6424. doi: 10.3934/mbe.2024280

| [1] |
E. Ranta, D. Tesar, V. Kaitala, Environmental variability and semelparity vs. iteroparity as life histories, J. Theor. Biol., 217 (2002), 391–396. https://doi.org/10.1006/jtb.2002.3029 doi: 10.1006/jtb.2002.3029

|
| [2] |
A. Margalida, M. Colomer, D. Oro, R. Arlettaz, J. Donázar, Assesing the impact of removal scenarios on population viability of a threatened long-lived avian scavenger, Sci. Rep., 5 (2015), 16962. https://doi.org/10.1038/srep16962 doi: 10.1038/srep16962
|
| [3] |
H. Tauler, J. Real, A. Hernández-Matías, P. Aymerich, J. Baucells, C. Martorell, et al., Identifying key demographic parameters for the viability of a growing population of the endangered Egyptian Vulture Neophron pernopterus, Bird Conserv. Int., 25 (2015), 246–439. https://doi.org/10.1017/S0959270914000392 doi: 10.1017/S0959270914000392
|
| [4] |
M. J. Faddy, Stochastic compartmental models as approximations to more general stochastic systems with the general stochastic epidemic as an example, Adv. Appl. Probab., 9 (1977), 448–461. https://doi.org/10.2307/1426108 doi: 10.2307/1426108
|
| [5] |
J. H. Matis, T. E. Wehrly, Stochastic models of compartmental systems, Biometrics, 35 (1979), 199–220. https://doi.org/10.2307/2529945 doi: 10.2307/2529945
|
| [6] | K. B. Athreya, P. E. Ney, Branching Processes, Springer-Verlag Berlin, Heidelberg, 1972. https://doi.org/10.1007/978-3-642-65371-1 |
| [7] | P. Guttorp, Statistical Inference for Branching Processes, Wiley, New York, 1991. |
| [8] | P. Haccou, P. Jagers, V. Vatutin, Branching Processes: Variation, Growth, and Extinction of Population, Cambridge University Press, Cambridge, 5 (2005). |
| [9] |
I. Pázsit, Symmetries and asymmetries in branching processes, Symmetry, 15 (2023), 1154. http://doi.org/10.3390/sym15061154 doi: 10.3390/sym15061154
|
| [10] |
L. A. Keating, J. P. Gleeson, D. J. O'Sullivan, Multitype branching process method for modeling complex contagion on clustered networks, Phys. Rev. E, 105 (2022), 034306. http://doi.org/10.1103/PhysRevE.105.034306 doi: 10.1103/PhysRevE.105.034306
|
| [11] | D. M. Hull, A survey of the literature associated with the bisexual Galton-Watson branching process, Extracta Math., 18 (2003), 321–343. |
| [12] | M. Molina, Two-sex branching process literature, in Workshop on Branching Processes and Their Applications, Springer Berlin Heidelberg, Berlin, 197 (2010), 279–293. https://doi.org/10.1007/978-3-642-11156-3-20 |
| [13] |
D. J. Daley, D. M. Hull, J. M. Taylor, Bisexual Galton-Watson branching processes with superadditive mating functions, J. Appl. Probab., 23 (1986), 585–600. https://doi.org/10.2307/3213999 doi: 10.2307/3213999
|
| [14] |
D. M. Hull, A necessary condition for extinction in those bisexual Galton-Watson branching processes governed for superadditive mating functions, J. Appl. Probab., 19 (1982), 847–850. https://doi.org/10.2307/3213838 doi: 10.2307/3213838
|
| [15] |
S. Ma, Y. Xing, The asymptotic properties of supercritical bisexual Galton-Watson branching processes with immigration of mating units, Acta Math. Sci., 26 (2006), 603–609. https://doi.org/10.1016/S0252-9602(06)60086-6 doi: 10.1016/S0252-9602(06)60086-6
|
| [16] |
M. Molina, C. Jacob, A. Ramos, Bisexual branching processes with offspring and mating depending on the number of couples in the population, Test, 17 (2008), 265–281. https://doi.org/10.1007/s11749-006-0031-9 doi: 10.1007/s11749-006-0031-9
|
| [17] |
Y. Xing, Y. Wang, On the extinction of one class of population-size dependent bisexual branching processes, J. Appl. Probab., 42 (2005), 175–184. https://doi.org/10.1239/jap/1110381379 doi: 10.1239/jap/1110381379
|
| [18] |
Y. Xing, Y. Wang, On the extinction of population-size dependent bisexual Galton-Watson processes, Acta Math. Sci., 42 (2008), 210–216. https://doi.org/10.1016/S0252-9602(08)60022-3 doi: 10.1016/S0252-9602(08)60022-3
|
| [19] |
M. Molina, M. Mota, A. Ramos, Stochastic modeling in biological populations with sexual reproduction through branching models: application to coho salmon populations, Math. Biosci., 258 (2014), 182–188. https://doi.org/10.1016/j.mbs.2014.10.007 doi: 10.1016/j.mbs.2014.10.007
|
| [20] |
M. Molina, M. Mota, A. Ramos, Two-sex branching processes with several mating and reproduction strategies: Extinction versus survival, Branching Processes Appl., 219 (2016), 307–317. https://doi.org/10.1007/978-3-319-31641-3-18 doi: 10.1007/978-3-319-31641-3-18
|
| [21] |
M. Molina, M. Mota, A. Ramos, Estimation of parameters in biological species with several mating and reproduction alternatives, Math. Biosci., 329 (2020), 108471. https://doi.org/10.1016/j.mbs.2020.108471 doi: 10.1016/j.mbs.2020.108471
|
| [22] | V. K. Rohatgi, An Introduction to Probability Theory and Mathematical Statistics, Wiley, New York, 1976. |
| [23] | B. W. Silverman, Density Estimation for Statistics and Data Analysis, Monographs on Statistics and Applied Probability, Chapman and Hall, London, 1986. |
| [24] | R Development Core Team, a Language and Enviroment for Statistical Computing, 2009. Available from: http://www.r-project.org. |
| [25] |
K. B. Karsten, L. N. Andriamandimbiarisoa, S. F. Fox, C. J. Raxworthy, A unique life history among tetrapods: An annual chameleon living mostly as an egg, Proc. Natl. Acad. Sci. USA, 105 (2008), 8980–8984. https://doi.org/10.1073/pnas.0802468105 doi: 10.1073/pnas.0802468105
|
| [26] |
F. Eckhardt, P. M. Kappeler, C. Kraus, Highly variable lifespan in an annual reptile, Labord's chameleon (Furcifer labordi), Sci. Rep., 7 (2017), 11397. https://doi.org/10.1038/s41598-017-11701-3 doi: 10.1038/s41598-017-11701-3
|
| [27] | F. Eckhardt, A Short Story: Senescence in an Annual Reptile, Labord's Cahameleon (Furcifer Labordi), Ph.D thesis, Georg-August University School of Science, 2019. Available from: http://dx.doi.org/10.53846/goediss-8355. |
| [28] |
L. Hudei, P. M. Kappeler, Sex-specific movement ecology of the shortest-lived tetrapod during the mating season, Sci. Rep., 12 (2022), 10053. https://doi.org/10.1038/s41598-022-14156-3 doi: 10.1038/s41598-022-14156-3
|
| [29] |
J. Ewald, P. Sieber, R. Garde, S. N. Lang, S. Schuster, B. Ibrahim, Trend in mathematical modeling of host-pathogen interactions, Cell. Mol. Life Sci., 77 (2020), 467–480. https://doi.org/10.1007/s00018-019-03382-0 doi: 10.1007/s00018-019-03382-0
|
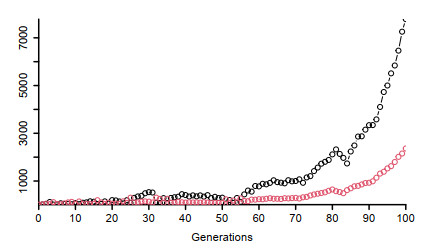
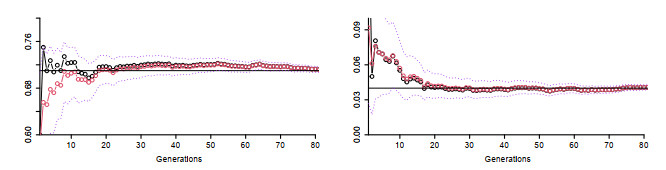
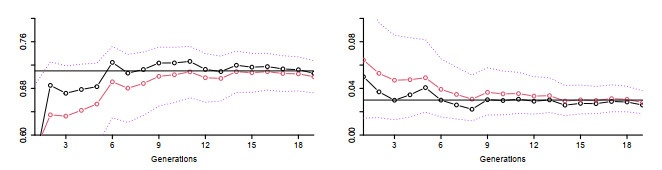
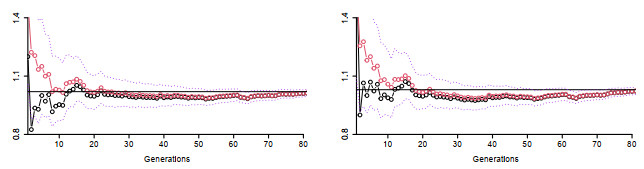
Figures(5) / Tables(2)

Manuel Molina, Manuel Mota, Alfonso Ramos. Mathematical modeling in semelparous biological species through two-sex branching processes[J]. Mathematical Biosciences and Engineering, 2024, 21(6): 6407-6424. doi: 10.3934/mbe.2024280







 DownLoad:
DownLoad: