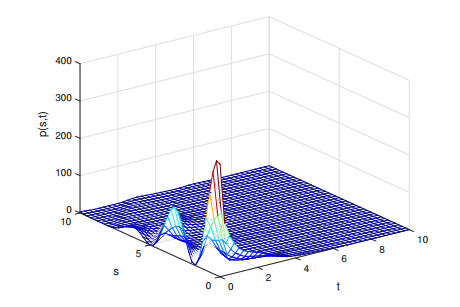
This paper investigates the existence of positive equilibrium as well as the stability of positive equilibrium and zero equilibrium in a nonlinear size-structured hierarchical population model. Under the condition that larger individuals are more competitive advantages than smaller ones, a non-zero fixed point theorem is used to show that there is at lest one positive equilibrium in the system. Moreover, we obtain the stability results of positive equilibrium and zero equilibrium by deriving characteristic equations and establishing Liapunov function. Finally, some numerical experiments are presented.
Citation: Weicheng Chen, Zhanping Wang. Stability of a class of nonlinear hierarchical size-structured population model[J]. Mathematical Biosciences and Engineering, 2022, 19(10): 10143-10159. doi: 10.3934/mbe.2022475
This paper investigates the existence of positive equilibrium as well as the stability of positive equilibrium and zero equilibrium in a nonlinear size-structured hierarchical population model. Under the condition that larger individuals are more competitive advantages than smaller ones, a non-zero fixed point theorem is used to show that there is at lest one positive equilibrium in the system. Moreover, we obtain the stability results of positive equilibrium and zero equilibrium by deriving characteristic equations and establishing Liapunov function. Finally, some numerical experiments are presented.

| [1] |
D. Dewsbury, Dominance rank, copulatory behavior, and differential reproduction, Q. Rev. Biol., 57 (1982), 135–159. https://doi.org/10.1086/412672 doi: 10.1086/412672

|
| [2] |
J. Cushing, The dynamics of hierarchical age-structured populations, J. Math. Biol., 32 (1994), 705–729. https://doi.org/10.1007/BF00163023 doi: 10.1007/BF00163023
|
| [3] | K. W. Blayneh, A hierarchical size-structured population model, Ph.D thesis, University of Arizona, 1996. |
| [4] |
À. Calsina, J. Saldaña, Asymptotic behaviour of a model of hierarchically structured population dynamics, J. Math. Biol., 35 (1997), 967–987. https://doi.org/10.1007/s002850050085 doi: 10.1007/s002850050085
|
| [5] |
S. Jang, J. Cushing, A discrete hierarchical model of intra-specific competition, J. Math. Anal. Appl., 280 (2003), 102–122. https://doi.org/10.1016/S0022-247X(03)00050-7 doi: 10.1016/S0022-247X(03)00050-7
|
| [6] |
A. S. Ackleh, K. Deng, J. Thibodeaux, A monotone approximation for a size-structured population model with a generalized environment, J. Biol. Dynam., 1 (2007), 305–319. https://doi.org/10.1080/17513750701605564 doi: 10.1080/17513750701605564
|
| [7] |
Y. Liu, Z. R. He, On the well-posedness of a nonlinear hierarchical size-structured population model, ANZIAM J., 58 (2017), 482–490. https://doi.org/10.21914/ANZIAMJ.V58I0.10831 doi: 10.21914/ANZIAMJ.V58I0.10831
|
| [8] |
A. S. Ackleh, K. Deng, S. H. Hu, A quasilinear hierarchical size-Structured model: well-posedness and approximation, Appl. Math. Opt., 51 (2005), 35–59. https://doi.org/10.1007/S00245-004-0806-2 doi: 10.1007/S00245-004-0806-2
|
| [9] |
E. Kraev, Existence and uniqueness for height structured hierarchical population model, Nat. Resour. Model., 14 (2008), 45–70. https://doi.org/10.1111/J.1939-7445.2001.TB00050.X doi: 10.1111/J.1939-7445.2001.TB00050.X
|
| [10] |
S. Henson, J. Cushing, Hierarchical models of intra-specific competition: Scramble versus contest, J. Math. Biol., 34 (1996), 755–772. https://doi.org/10.1007/BF00161518 doi: 10.1007/BF00161518
|
| [11] |
J. Cushing, A size-structured model for cannibalism, Theor. Popul. Biol., 42 (1992), 347–361. https://doi.org/10.1016/0040-5809(92)90020-T doi: 10.1016/0040-5809(92)90020-T
|
| [12] |
W. Gurney, R. Nisbet, Ecological stability and social hierarchy, Theor. Popul. Biol., 16 (1979), 48–80. https://doi.org/10.1016/0040-5809(79)90006-6 doi: 10.1016/0040-5809(79)90006-6
|
| [13] |
J. Z. Farkas, T. Hagen, Stability and regularity results for a size-structured population model, J. Math. Anal. Appl., 328 (2007), 119–136. https://doi.org/10.1016/J.JMAA.2006.05.032 doi: 10.1016/J.JMAA.2006.05.032
|
| [14] |
X. R. Li, The stability of nonlinear age-dependent population equation, Appl. Math. Lett., 11 (1998), 19–26. https://doi.org/10.1016/S0893-9659(98)00096-2 doi: 10.1016/S0893-9659(98)00096-2
|
| [15] |
J. Z. Farkas, P. Hinow, Steady states in hierarchical structured populations with distributed states at birth, Discrete Contin. Dyn. Syst. B, 17 (2012), 2671–2689. https://doi.org/10.3934/dcdsb.2012.17.2671 doi: 10.3934/dcdsb.2012.17.2671
|
| [16] | Z. R. He, Z. Q. Zhang, Y. Wang, Stability of a class of nonlinear hierarchical age-dependent population model (in Chinese), Acta Math. Sci. Ser. A, 40 (2020), 1712–1722. http://121.43.60.238/sxwlxbA/CN/Y2020/V40/I6/1712 |
| [17] |
Z. R. He, N. Zhou, Stability for a competing system of hierarchical age-structured populations, Int. J. Biomath., 13 (2020), 2050070. https://doi.org/10.1142/S1793524520500709 doi: 10.1142/S1793524520500709
|
| [18] |
J. Prüss, On the qualitative behaviour of populations with age-specific interactions, Comput. Math. Appl., 9 (1983), 327–339. https://doi.org/10.1016/0898-1221(83)90020-2 doi: 10.1016/0898-1221(83)90020-2
|
| [19] |
M. Farkas, On the stability of stationary age distributions, Appl. Math. Comput., 131 (2002), 107–123. https://doi.org/10.1016/S0096-3003(01)00131-X doi: 10.1016/S0096-3003(01)00131-X
|
| [20] |
J. Z. Farkas, Stability conditions for a non-linear size-structured model, Nonlinear Anal. Real World Appl., 6 (2006), 962–969. https://doi.org/10.1016/J.NONRWA.2004.06.002 doi: 10.1016/J.NONRWA.2004.06.002
|
| [21] | K. Yosida, Function Analysis, 6th edition, Springer-Verlag, Berlin, 1980. https://doi.org/10.1007/978-3-662-25762-3 |





Figures(6)

Weicheng Chen, Zhanping Wang. Stability of a class of nonlinear hierarchical size-structured population model[J]. Mathematical Biosciences and Engineering, 2022, 19(10): 10143-10159. doi: 10.3934/mbe.2022475







 DownLoad:
DownLoad: