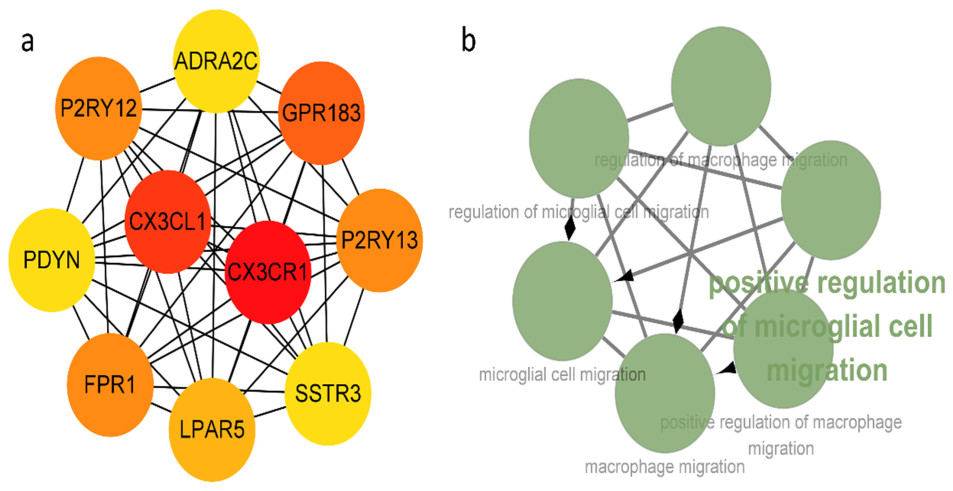
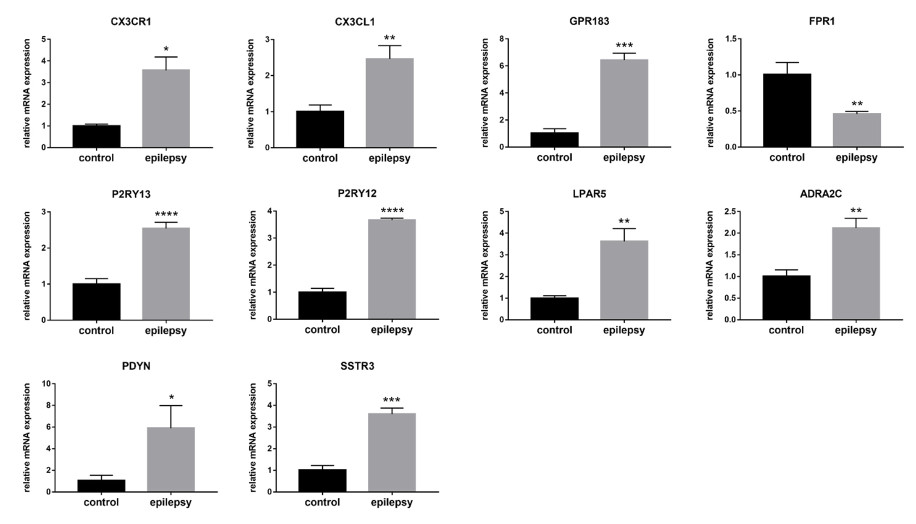
Several studies have shown a link between immunity, inflammatory processes, and epilepsy. Active neuroinflammation and marked immune cell infiltration occur in epilepsy of diverse etiologies. Microglia, as the first line of defense in the central nervous system, are the main effectors of neuroinflammatory processes. Discovery of new biomarkers associated with microglia activation after epileptogenesis indicates that targeting specific molecules may help control seizures. In this research, we used a combination of several bioinformatics approaches, including RNA sequencing, to explore differentially expressed genes (DEGs) in epileptic lesions and control samples, and to construct a protein-protein interaction (PPI) network for DEGs, which was examined utilizing plug-ins in Cytoscape software. Finally, we aimed to identify 10 hub genes in immune and inflammation-related sub-networks, which were subsequently validated in real-time quantitative polymerase chain reaction analysis in a mouse model of kainic acid-induced epilepsy. The expression patterns of nine genes were consistent with sequencing outcomes. Meanwhile, several genes, including CX3CR1, CX3CL1, GPR183, FPR1, P2RY13, P2RY12 and LPAR5, were associated with microglial activation and migration, providing novel candidate targets for immunotherapy in epilepsy and laying the foundation for further research.
Citation: Jing Cao, Hui Gan, Han Xiao, Hui Chen, Dan Jian, Ning Jiang, Xuan Zhai. Key protein-coding genes related to microglia in immune regulation and inflammatory response induced by epilepsy[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 9563-9578. doi: 10.3934/mbe.2021469
Several studies have shown a link between immunity, inflammatory processes, and epilepsy. Active neuroinflammation and marked immune cell infiltration occur in epilepsy of diverse etiologies. Microglia, as the first line of defense in the central nervous system, are the main effectors of neuroinflammatory processes. Discovery of new biomarkers associated with microglia activation after epileptogenesis indicates that targeting specific molecules may help control seizures. In this research, we used a combination of several bioinformatics approaches, including RNA sequencing, to explore differentially expressed genes (DEGs) in epileptic lesions and control samples, and to construct a protein-protein interaction (PPI) network for DEGs, which was examined utilizing plug-ins in Cytoscape software. Finally, we aimed to identify 10 hub genes in immune and inflammation-related sub-networks, which were subsequently validated in real-time quantitative polymerase chain reaction analysis in a mouse model of kainic acid-induced epilepsy. The expression patterns of nine genes were consistent with sequencing outcomes. Meanwhile, several genes, including CX3CR1, CX3CL1, GPR183, FPR1, P2RY13, P2RY12 and LPAR5, were associated with microglial activation and migration, providing novel candidate targets for immunotherapy in epilepsy and laying the foundation for further research.

| [1] | O. Nevalainen, H. Ansakorpi, M. Simola, J. Raitanen, J. Isojärvi, M. Artama, et al., Epilepsy-related clinical characteristics and mortality: a systematic review and meta-analysis, Neurology, 83 (2014), 1968–1977. |
| [2] | S. Popkirov, F. S. Ismail, W. Grönheit, M. Kapauer, J. Wellmer, C. G. Bien, Progressive hippocampal sclerosis after viral encephalitis: Potential role of NMDA receptor antibodies, Seizure, 51 (2017), 6–8. |
| [3] | J. Choi, D. R. Nordli, T. D. Alden, A. DiPatri, L. Laux, K. Kelley, et al., Cellular injury and neuroinflammation in children with chronic intractable epilepsy, J. Neuroinflammation, 6 (2009), 38. |
| [4] | A. Vezzani, S. Balosso, T. Ravizza, The role of cytokines in the pathophysiology of epilepsy, Brain Behav. Immun., 22 (2008), 797–803. |
| [5] | T. Ravizza, B. Gagliardi, F. Noé, K. Boer, E. Aronica, A. Vezzani, Innate and adaptive immunity during epileptogenesis and spontaneous seizures: evidence from experimental models and human temporal lobe epilepsy, Neurobiol. Dis., 29 (2008), 142–160. |
| [6] | K. Rijkers, H. J. Majoie, G. Hoogland, G. Kenis, M. D. Baets, J. S. Vles, The role of interleukin-1 in seizures and epilepsy: a critical review, Exp. Neurol., 216 (2009), 258–271. |
| [7] | M. M. Nagib, Y. Yu, J. Jiang, Targeting prostaglandin receptor EP2 for adjunctive treatment of status epilepticus, Pharmacol. Ther., 209 (2020), 107504. |
| [8] | A. Dey, X. Kang, J. Qiu, Y. Du, J. Jiang, Anti-inflammatory small molecules to treat seizures and epilepsy: from bench to bedside, Trends Pharmacol. Sci., 37 (2016), 463–484. |
| [9] | K. Boer, W. G. Spliet, P. C. V. Rijen, S. Redeker, D. Troost, E. Aronica, Evidence of activated microglia in focal cortical dysplasia, J. Neuroimmunol., 173 (2006), 188–195. |
| [10] | M. J. Benson, S. Manzanero, K. Borges, Complex alterations in microglial M1/M2 markers during the development of epilepsy in two mouse models, Epilepsia, 56 (2015), 895–905. |
| [11] | G. Kalozoumi, O. Kel-Margoulis, E. Vafiadaki, D. Greenberg, H. Bernard, H. Soreq, et al., Glial responses during epileptogenesis in Mus musculus point to potential therapeutic targets, PLoS One, 13 (2018), e0201742. |
| [12] | Z. H. Luo, A. A. Walid, Y. Xie, H. Long, W. Xiao, L. Xu, et al., Construction and analysis of a dysregulated lncRNA-associated ceRNA network in a rat model of temporal lobe epilepsy, Seizure, 69 (2019), 105–114. |
| [13] | Q. L. Chen, L. Xia, S. P. Zhong, Q. Wang, J. Ding, Bioinformatic analysis identifies key transcriptome signatures in temporal lobe epilepsy, Neuroscience, 26 (2020), 1266–1277. |
| [14] | V. Riban, V. Bouilleret, B. T. Pham-Lê, J. M. Fritschy, C. Marescaux, A. Depaulis, Evolution of hippocampal epileptic activity during the development of hippocampal sclerosis in a mouse model of temporal lobe epilepsy, Neuroscience, 112 (2002), 101–111. |
| [15] | A. Mortazavi, B. A. Williams, K. McCue, L. Schaeffer, B. Wold, Mapping and quantifying mammalian transcriptomes by RNA-Seq, Nat. Methods, 5 (2008), 621–628. |
| [16] | G. D. Bader, C. W. Hogue, An automated method for finding molecular complexes in large protein interaction networks, BMC Bioinf., 4 (2003), 2. |
| [17] | C. H. Chin, S. H. Chen, H. H. Wu, C. W. Ho, M. T. Ko, C. Y. Lin, cytoHubba: identifying hub objects and sub-networks from complex interactome, BMC Syst. Biol., 8 (2014), S11. |
| [18] | G. Bindea, B. Mlecnik, H. Hackl, P. Charoentong, M. Tosolini, A. Kirilovsky, et al., ClueGO: a Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks, Bioinformatics, 25 (2009), 1091–1093. |
| [19] | G. Bindea, J. Galon, B. Mlecnik, CluePedia Cytoscape plugin: pathway insights using integrated experimental and in silico data, Bioinformatics, 29 (2013), 661–663. |
| [20] | X. He, J. Zhang, Why do hubs tend to be essential in protein networks?, PLoS Genet., 2 (2006), e88. |
| [21] | N. Yu, H. Liu, Q. Di, Modulation of Immunity and the Inflammatory Response: A New Target for Treating Drug-resistant Epilepsy, Curr. Neuropharmacol., 11 (2013), 114–127. |
| [22] | S. Kinoshita, R. Koyama, Pro- and anti-epileptic roles of microglia, Neural. Regen. Res., 16 (2021), 1369–1371. |
| [23] | A. Nishiyori, M. Minami, Y. Ohtani, S. Takami, J. Yamamoto, N. Kawaguchi, et al., Localization of fractalkine and CX3CR1 mRNAs in rat brain: does fractalkine play a role in signaling from neuron to microglia?, FEBS Lett., 429 (1998), 167–172. |
| [24] | J. K. Harrison, Y. Jiang, S. Chen, Y. Xia, D. Maciejewski, R. K. McNamara, et al., Role for neuronally derived fractalkine in mediating interactions between neurons and CX3CR1-expressing microglia, Proc. Nat. Acad. Sci. U. S. A., 95 (1998), 10896–10901. |
| [25] | O. Meucci, A. Fatatis, A. A. Simen, R. J. Miller, Expression of CX3CR1 chemokine receptors on neurons and their role in neuronal survival, Proc. Nat. Acad. Sci. U. S. A., 97 (2000), 8075–8080. |
| [26] | A. E. Cardona, E. P. Pioro, M. E. Sasse, V. Kostenko, S. M. Cardona, I. M. Dijkstra, et al., Control of microglial neurotoxicity by the fractalkine receptor, Nat. Neurosci., 9 (2006), 917–924. |
| [27] | A. D. Bachstetter, J. M. Morganti, J. Jernberg, A. Schlunk, S. H. Mitchell, K. W. Brewster, et al., Fractalkine and CX 3 CR1 regulate hippocampal neurogenesis in adult and aged rats, Neurobiol. Aging, 32 (2011), 2030–2044. |
| [28] | Y. Xu, K. Zeng, Y. Han, L. Wang, D. Chen, Z. Xi, et al., Altered expression of CX3CL1 in patients with epilepsy and in a rat model, Am. J. Pathol., 180 (2012), 1950–1962. |
| [29] | C. Roseti, S. Fucile, C. Lauro, K. Martinello, C. Bertollini, V. Esposito, et al., Fractalkine/CX3CL1 modulates GABAA currents in human temporal lobe epilepsy, Epilepsia, 54 (2013), 1834–1844. |
| [30] | U. B. Eyo, J. Peng, M. Murugan, M. Mo, A. Lalani, P. Xie, et al., Regulation of physical microglia-neuron interactions by fractalkine signaling after status epilepticus, eNeuro, 3 (2016). |
| [31] | S. Hannedouche, J. Zhang, T. Yi, W. Shen, D. Nguyen, J. P. Pereira, et al., Oxysterols direct immune cell migration via EBI2, Nature, 475 (2011), 524–527. |
| [32] | A. Rutkowska, I. Preuss, F. Gessier, A. W. Sailer, K. K. Dev, EBI2 regulates intracellular signaling and migration in human astrocyte, Glia, 63 (2015), 341–351. |
| [33] | F. Wanke, S. Moos, A. L. Croxford, A. P. Heinen, S. Gräf, B. Kalt, et al., EBI2 is highly expressed in multiple sclerosis lesions and promotes early CNS migration of encephalitogenic CD4 T cells, Cell Rep., 18 (2017), 1270–1284. |
| [34] | I. Klejbor, D. R. Shimshek, J. Klimaszewska-Łata, M. Velasco-Estevez, J. Moryś, B. Karaszewski, et al., EBI2 is expressed in glial cells in multiple sclerosis lesions, and its knock-out modulates remyelination in the cuprizone model, Eur. J. Neurosci., (2021). |
| [35] | F. Boulay, M. Tardif, L. Brouchon, P. Vignais, Synthesis and use of a novel N-formyl peptide derivative to isolate a human N-formyl peptide receptor cDNA, Biochem. Biophys. Res. Commun., 168 (1990), 1103–1109. |
| [36] | D. C. Maestes, R. M. Potter, E. R. Prossnitz, Differential phosphorylation paradigms dictate desensitization and internalization of the N-formyl peptide receptor, J. Biol. Chem., 274 (1999), 29791–29795. |
| [37] | B. Postma, M. J. Poppelier, J. C. V. Galen, E. R. Prossnitz, J. A. V. Strijp, C. J. de Haas, et al., Chemotaxis inhibitory protein of Staphylococcus aureus binds specifically to the C5a and formylated peptide receptor, J. Immunol., 172 (2004), 6994–7001. |
| [38] | K. Bihler, E. Kress, S. Esser, S. Nyamoya, S. C. Tauber, T. Clarner, et al., Formyl peptide receptor 1-mediated glial cell activation in a mouse model of cuprizone-induced demyelination, J. Mol. Neurosci., 62 (2017), 232–243. |
| [39] | G. Wang, L. Zhang, X. Chen, X. Xue, Q. Guo, M. Liu, et al., Formylpeptide receptors promote the migration and differentiation of rat neural stem cells, Sci. Rep., 6 (2016), 25946. |
| [40] | Y. Sasaki, M. Hoshi, C. Akazawa, Y. Nakamura, H. Tsuzuki, K. Inoue, et al., Selective expression of Gi/o-coupled ATP receptor P2Y12 in microglia in rat brain, Glia, 44 (2003), 242–250. |
| [41] | T. R. Hammond, C. Dufort, L. Dissing-Olesen, S. Giera, A. Young, A. Wysoker, et al., Single-cell RNA sequencing of microglia throughout the mouse lifespan and in the injured brain reveals complex cell-state changes, Immunity, 50 (2019), 253–271. |
| [42] | U. B. Eyo, M. Mo, M. H. Yi, M. Murugan, J. Liu, R. Yarlagadda, et al., P2Y12R-dependent translocation mechanisms gate the changing microglial landscape, Cell Rep., 23 (2018), 959–966. |
| [43] | S. E. Haynes, G. Hollopeter, G. Yang, D. Kurpius, M. E. Dailey, W. B. Gan, et al., The P2Y12 receptor regulates microglial activation by extracellular nucleotides, Nat. Neurosci., 9 (2006), 1512–1519. |
| [44] | U. B. Eyo, J. Peng, P. Swiatkowski, A. Mukherjee, A. Bispo, L. J. Wu, Neuronal hyperactivity recruits microglial processes via neuronal NMDA receptors and microglial P2Y12 receptors after status epilepticus, J. Neurosci., 34 (2014), 10528–10540. |
| [45] | M. Mo, U. B. Eyo, M. Xie, J. Peng, D. B. Bosco, A. D. Umpierre, et al., Microglial P2Y12 receptor regulates seizure-induced neurogenesis and immature neuronal projections, J. Neurosci., 39 (2019), 9453–9464. |
| [46] | K. Kobayashi, H. Yamanaka, F. Yanamoto, M. Okubo, K. Noguchi, Multiple P2Y subtypes in spinal microglia are involved in neuropathic pain after peripheral nerve injury, Glia, 60 (2012), 1529–1539. |
| [47] | V. Morente, R. Pérez-Sen, F. Ortega, J. Huerta-Cepas, E. G. Delicado, M. T. Miras-Portugal, Neuroprotection elicited by P2Y13 receptors against genotoxic stress by inducing DUSP2 expression and MAPK signaling recovery, Biochem. Biophys. Acta, 1843 (2014), 1886–1898. |
| [48] | V. Kyrargyri, C. Madry, A. Rifat, I. L. Arancibia-Carcamo, S. P. Jones, V. T. T. Chan, et al., P2Y(13) receptors regulate microglial morphology, surveillance, and resting levels of interleukin 1β release, Glia, 68 (2020), 328–344. |
| [49] | P. Jiang, F. Xing, B. Guo, J. Yang, Z. Li, W. Wei, et al., Nucleotide transmitters ATP and ADP mediate intercellular calcium wave communication via P2Y12/13 receptors among BV-2 microglia, PLoS One, 12 (2017), e0183114. |
| [50] | M. E. Lin, R. R. Rivera, J. Chun, Targeted deletion of LPA5 identifies novel roles for lysophosphatidic acid signaling in development of neuropathic pain, J. Biol. Chem., 287 (2012), 17608–17617. |
| [51] | R. Tsukahara, S. Yamamoto, K. Yoshikawa, M. Gotoh, T. Tsukahara, H. Neyama, et al., LPA5 signaling is involved in multiple sclerosis-mediated neuropathic pain in the cuprizone mouse model, J. Pharmacol. Sci., 136 (2018), 93–96. |
| [52] | I. Plastira, E. Bernhart, M. Goeritzer, H. Reicher, V. B. Kumble, N. Kogelnik, et al., 1-Oleyl-lysophosphatidic acid (LPA) promotes polarization of BV-2 and primary murine microglia towards an M1-like phenotype, J. Neuroinflammation, 13 (2016), 205. |
| [53] | I. Plastira, E. Bernhart, M. Goeritzer, T. DeVaney, H. Reicher, A. Hammer, et al., Lysophosphatidic acid via LPA-receptor 5/protein kinase D-dependent pathways induces a motile and pro-inflammatory microglial phenotype, J. Neuroinflammation, 14 (2017), 253. |
| [54] | I. Plastira, L. Joshi, E. Bernhart, J. Schoene, E. Specker, M. Nazare, et al., Small-molecule lysophosphatidic acid receptor 5 (LPAR5) antagonists: versatile pharmacological tools to regulate inflammatory signaling in BV-2 microglia cells, Front. Cell Neurosci., 13 (2019), 531. |
| [55] | L. Hein, J. D. Altman, B. K. Kobilka, Two functionally distinct alpha2-adrenergic receptors regulate sympathetic neurotransmission, Nature, 402 (1999), 181–184. |
| [56] | C. Chavkin, Dynorphin--still an extraordinarily potent opioid peptide, Mol. Pharmacol., 83 (2013), 729–736. |
| [57] | M. Simonato, P. Romualdi, Dynorphin and epilepsy, Prog. Neurobiol., 50 (1996), 557–583. |
| [58] | S. Loacker, M. Sayyah, W. Wittmann, H. Herzog, C. Schwarzer, Endogenous dynorphin in epileptogenesis and epilepsy: anticonvulsant net effect via kappa opioid receptors, Brain, 130 (2007), 1017–1028. |
| [59] | B. Queenan, R. Dunn, V. Santos, Y. Feng, M. Huizenga, R. Hammack, et al., Kappa opioid receptors regulate hippocampal synaptic homeostasis and epileptogenesis, Epilepsia, 59 (2018), 106–122. |
| [60] | H. Dai, P. Wang, H. Mao, X. Mao, S. Tan, Z. Chen, Dynorphin activation of kappa opioid receptor protects against epilepsy and seizure-induced brain injury via PI3K/Akt/Nrf2/HO-1 pathway, Cell Cycle, 18 (2019), 226–237. |
| [61] | L. Liu, Y. Xu, H. Dai, S. Tan, X. Mao, Z. Chen, Dynorphin activation of kappa opioid receptor promotes microglial polarization toward M2 phenotype via TLR4/NF-κB pathway, Cell Biosci., 10 (2020), 42. |
| [62] | L. Zangrandi, C. Schwarzer, The kappa opioid receptor system in temporal lobe epilepsy, Handb. Exp. Pharmacol., 2021. |
| [63] | D. Stanić, H. Malmgren, H. He, L. Scott, A. Aperia, T. Hökfelt, Developmental changes in frequency of the ciliary somatostatin receptor 3 protein, Brain Res., 1249 (2009), 101–112. |
| [64] | Q. Yao, Q. Liu, H. Xu, Z. Wu, L. Zhou, Z. Gu, et al., Upregulated expression of SSTR3 is involved in neuronal apoptosis after intracerebral hemorrhage in adult rats, Cell Mol. Neurobiol., 37 (2017), 1407–1416. |
| [65] | S. Nielsen, S. Mellemkjaer, L. M. Rasmussen, T. Ledet, N. Olsen, M. Bojsen-Møller, et al., Expression of somatostatin receptors on human pituitary adenomas in vivo and ex vivo, J. Endocrinol. Invest., 24 (2001), 430–437. |
| [66] | R. Panetta, Y. C. Patel, Expression of mRNA for all five human somatostatin receptors (hSSTR1-5) in pituitary tumors, Life Sci., 56 (1995), 333–342. |
| [67] | K. Lamszus, W. Meyerhof, M. Westphal, Somatostatin and somatostatin receptors in the diagnosis and treatment of gliomas, J. Neurooncol., 35 (1997), 353–364. |
| [68] | H. Luo, C. Tao, X. Long, K. Huang, X. Zhu, A risk signature of four aging-related genes has clinical prognostic value and is associated with a tumor immune microenvironment in glioma, Aging (Albany NY) 13 (2021), 16198–16218. |
Figures(5) / Tables(4)

Jing Cao, Hui Gan, Han Xiao, Hui Chen, Dan Jian, Ning Jiang, Xuan Zhai. Key protein-coding genes related to microglia in immune regulation and inflammatory response induced by epilepsy[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 9563-9578. doi: 10.3934/mbe.2021469







 DownLoad:
DownLoad: