Citation: Hwayeon Ryu, Sue Ann Campbell. Stability, bifurcation and phase-locking of time-delayed excitatory-inhibitory neural networks[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7931-7957. doi: 10.3934/mbe.2020403

| [1] | P. Dayan, L. Abbott, Theoretical Neuroscience, MIT Press, Cambridge, MA, 2001. |
| [2] | G. Ermentrout, D. Terman, Mathematical Foundations of Neuroscience, Springer, New York, NY, 2010. |
| [3] | E. M. Izhikevich, Dynamical Systems in Neuroscience, MIT Press, Cambridge, MA, 2007. |
| [4] | N. Brunel, X. J. Wang, What determines the frequency of fast network oscillations with irregular neural discharges? I. synaptic dynamics and excitation-inhibition balance, J. Neurophysiol., 90 (2003), 415-430. |
| [5] |
G. B. Ermentrout, N. Kopell, Fine structure of neural spiking and synchronization in the presence of conduction delays, Proc. Natl. Acad. Sci., 95 (1998), 1259-1264. doi: 10.1073/pnas.95.3.1259

|
| [6] |
N. Kopell, G. Ermentrout, M. Whittington, R. Traub, Gamma rhythms and beta rhythms have different synchronization properties, Proc. Natl. Acad. Sci., 97 (2000), 1867-1872. doi: 10.1073/pnas.97.4.1867
|
| [7] |
J. E. Rubin, D. Terman, Analysis of clustered firing patterns in synaptically coupled networks of oscillators, J. Math. Biol., 41 (2000), 513-545. doi: 10.1007/s002850000065
|
| [8] |
B. Pfeuty, G. Mato, D. Golomb, D. Hansel, The combined effects of inhibitory and electrical synapses in synchrony, Neural Comput., 17 (2005), 633-670. doi: 10.1162/0899766053019917
|
| [9] |
M. A. Whittington, R. Traub, N. Kopell, B. Ermentrout, E. Buhl, Inhibition-based rhythms: experimental and mathematical observations on network dynamics, Int. J. Psychophysiol., 38 (2000), 315-336. doi: 10.1016/S0167-8760(00)00173-2
|
| [10] |
J. E. Rubin, D. Terman, Geometric analysis of population rhythms in synaptically coupled neuronal networks, Neural Comput., 12 (2000), 597-645. doi: 10.1162/089976600300015727
|
| [11] | J. E. Rubin, D. Terman, Geometric singular perturbation analysis of neuronal dynamics, Handbook Dyn. Syst., 2 (2002), 93-146. |
| [12] |
C. D. Acker, N. Kopell, J. A. White, Synchronization of strongly coupled excitatory neurons: relating network behavior to biophysics, J. Comput. Neurosci., 15 (2003), 71-90. doi: 10.1023/A:1024474819512
|
| [13] | P. J. Hellyer, B. Jachs, C. Clopath, R. Leech, Local inhibitory plasticity tunes macroscopic brain dynamics and allows the emergence of functional brain networks, Neuroimage, 24 (2016), 85-95. |
| [14] |
E. M. Izhikevich, G. M. Edelman, Large-scale model of mammalian thalamocortical systems, Proc. Natl. Acad. Sci., 105 (2008), 3593-3598. doi: 10.1073/pnas.0707563105
|
| [15] |
M. Bazhenov, N. F. Rulkov, I. Timofeev, Effect of synaptic connectivity on long-range synchronization of fast cortical oscillations, J. Neurophysiol., 100 (2008), 1562-1575. doi: 10.1152/jn.90613.2008
|
| [16] |
A. Ghosh, Y. Rho, A. McIntosh, R. Kötter, V. Jirsa, Cortical network dynamics with time delays reveals functional connectivity in the resting brain, Cognitive Neurodyn., 2 (2008), 115. doi: 10.1007/s11571-008-9044-2
|
| [17] |
D. Guo, Q. Wang, M. Perc, Complex synchronous behavior in interneuronal networks with delayed inhibitory and fast electrical synapses, Phys. Rev. E, 85 (2012), 061905. doi: 10.1103/PhysRevE.85.061905
|
| [18] |
T. Pérez, G. C. Garcia, V. M. Eguiluz, R. Vicente, G. Pipa, C. Mirasso, Effect of the topology and delayed interactions in neuronal networks synchronization, PLoS ONE, 6 (2011), e19900. doi: 10.1371/journal.pone.0019900
|
| [19] | X. Sun, L. Guofang, Fast regular firings induced by intra- and inter-time delays in two clustered neuronal networks, Chaos, 28 (2017), 106310. |
| [20] |
Q. Wang, Q. Lu, G. Chen, Synchronization transition induced by synaptic delay in coupled fastspiking neurons, Int. J. Bifurcat. Chaos, 18 (2008), 1189-1198. doi: 10.1142/S0218127408020914
|
| [21] |
S. Crook, G. Ermentrout, M. Vanier, J. Bower, The role of axonal delay in synchronization of networks of coupled cortical oscillators, J. Comput. Neurosci., 4 (1997), 161-172. doi: 10.1023/A:1008843412952
|
| [22] |
P. Bressloff, S. Coombes, Symmetry and phase-locking in a ring of pulse-coupled oscillators with distributed delays, Physica D, 126 (1999), 99-122. doi: 10.1016/S0167-2789(98)00264-4
|
| [23] |
P. Bressloff, S. Coombes, Travelling waves in chains of pulse-coupled integrate-and-fire oscillators with distributed delays, Physica D, 130 (1999), 232-254. doi: 10.1016/S0167-2789(99)00013-5
|
| [24] |
T. W. Ko, G. B. Ermentrout, Effects of axonal time delay on synchronization and wave formation in sparsely coupled neuronal oscillators, Phys. Rev. E, 76 (2007), 056206. doi: 10.1103/PhysRevE.76.056206
|
| [25] |
S. A. Campbell, Z. Wang, Phase models and clustering in networks of oscillators with delayed coupling, Physica D, 363 (2018), 44-55. doi: 10.1016/j.physd.2017.09.004
|
| [26] |
G. Orosz, Decomposition of nonlinear delayed networks around cluster states with applications to neurodynamics, SIAM J. Appl. Dyn. Syst., 13 (2014), 1353-1386. doi: 10.1137/130915637
|
| [27] |
Z. Wang, S. A. Campbell, Symmetry, Hopf bifurcation, and the emergence of cluster solutions in time delayed neural networks, Chaos, 27 (2017), 114316. doi: 10.1063/1.5006921
|
| [28] |
D. Golomb, G. B. Ermentrout, Continuous and lurching traveling pulses in neuronal networks with delay and spatially decaying connectivity, Proc. Natl. Acad. Sci., 96 (1999), 13480-13485. doi: 10.1073/pnas.96.23.13480
|
| [29] |
D. Golomb, G. B. Ermentrout, Effects of delay on the type and velocity of travelling pulses in neuronal networks with spatially decaying connectivity, Network: Comput. Neural Syst., 11 (2000), 221-246. doi: 10.1088/0954-898X_11_3_304
|
| [30] |
A. Roxin, N. Brunel, D. Hansel, Role of delays in shaping spatiotemporal dynamics of neuronal activity in large networks, Phys. Rev. Lett.s, 94 (2005), 238103. doi: 10.1103/PhysRevLett.94.238103
|
| [31] | M. Dahlem, G. Hiller, A. Panchuk, E. Schöll, Dynamics of delay-coupled excitable neural systems, Int. J. Bifurc. Chaos, 29 (2009), 745-753. |
| [32] |
N. Burić, I. Grozdanović, N. Vasović, Type I vs type II excitable systems with delayed coupling, Chaos Solitons Fractals, 23 (2005), 1221-1233. doi: 10.1016/j.chaos.2004.06.033
|
| [33] | N. Burić, D. Todorović, Dynamics of Fitzhugh-Nagumo excitable systems with delayed coupling, Phys. Rev. E, 67 (2003), 066-222. |
| [34] |
N. Burić, D. Todorović, Bifurcations due to small time-lag in coupled excitable systems, Int. J. Bifurc. Chaos, 15 (2005), 1775-1785. doi: 10.1142/S0218127405012831
|
| [35] | S. A. Campbell, R. Edwards, P. van den Dreissche, Delayed coupling between two neural network loops, SIAM J. Appl. Math., 65 (2004), 316-335. |
| [36] |
E. Schöll, G. Hiller, P. Hövel, M. A. Dahlem, Time-delay feedback in neurosystems, Phil. Trans. Roy. Soc. A, 367 (2009), 1079-10956. doi: 10.1098/rsta.2008.0258
|
| [37] |
C. U. Choe, T. Dahms, P. Hovel, E. Schöll, Controlling synchrony by delay coupling in networks: From in-phase to splay and cluster states, Phys. Rev. E, 81 (2010), 025205. doi: 10.1103/PhysRevE.81.025205
|
| [38] |
T. Dahms, J. Lehnert, E. Scholl, Cluster and group synchronization in delay-coupled networks, Phys. Rev. E, 86 (2012), 016202. doi: 10.1103/PhysRevE.86.016202
|
| [39] |
J. Lehnert, T. Dahms, P. Hovel, E. Schöll, Loss of synchronization in complex neuronal networks with delay, EPL (Europhys. Lett.), 96 (2011), 60013. doi: 10.1209/0295-5075/96/60013
|
| [40] |
Y. Song, J. Xu, Inphase and antiphase synchronization in a delay-coupled system with applications to a delay-coupled Fitzhugh-Nagumo system, IEEE Trans. Neural. Netw. Learn. Syst., 23 (2012), 1659-1670. doi: 10.1109/TNNLS.2012.2209459
|
| [41] | A. Panchuk, D. Rosin, P. Hovel, E. Schöll, Synchronization of coupled neural oscillators with heterogeneous delays, Int. J. Bifurc. Chaos, 23. |
| [42] |
S. R. Campbell, D. Wang, Relaxation oscillators with time delay coupling, Physica D, 111 (1998), 151-178. doi: 10.1016/S0167-2789(97)80010-3
|
| [43] |
J. J. Fox, C. Jayaprakash, D. Wang, S. R. Campbell, Synchronization in relaxation oscillator networks with conduction delays, Neural Comput., 13 (2001), 1003-1021. doi: 10.1162/08997660151134307
|
| [44] | A. Bose, S. Kunec, Synchrony and frequency regulation by synaptic delay in networks of selfinhibiting neurons, Neurocomputing, 38 (2001), 505-513. |
| [45] |
S. Kunec, A. Bose, Role of synaptic delay in organizing the behavior of networks of self-inhibiting neurons, Phys. Rev. E, 63 (2001), 021908. doi: 10.1103/PhysRevE.63.021908
|
| [46] |
S. Kunec, A. Bose, High-frequency, depressing inhibition facilitates synchronization in globally inhibitory networks, Network: Comput. Neural Syst., 14 (2003), 647-672. doi: 10.1088/0954-898X_14_4_303
|
| [47] | H. Ryu, S. A. Campbell, Geometric analysis of synchronization in neuronal networks with global inhibition and coupling delays, Phil. Trans. R. Soc. A, 337 (2019), 20180129. |
| [48] | X. Ji, J. Lu, J. Lou, J. Qiu, K. Shi, A unified criterion for global exponential stability of quaternionvalued neural networks with hybrid impulses, Int. J. Robust Nonlinear Control, 2020. |
| [49] | B. Sun, Y. Cao, Z. Guo, Z. Yan, S. Wen, Synchronization of discrete-time recurrent neural networks with time-varying delays via quantized sliding mode control, Appl. Math. Comput., 375 (2020), 125093. |
| [50] |
Y. Tian, Z. Wang, Stability analysis for delayed neural networks based on the augmented Lyapunov-Krasovskii functional with delay-product-type and multiple integral terms, Neurocomputing, 410 (2020), 295-303. doi: 10.1016/j.neucom.2020.05.045
|
| [51] |
J. Xiao, Z. Zeng, A. Wu, S. Wen, Fixed-time synchronization of delayed Cohen-Grossberg neural networks based on a novel sliding mode, Neural Networks, 128 (2020), 1-12. doi: 10.1016/j.neunet.2020.04.020
|
| [52] | E. Ermentrout, Simulating, Analyzing, and Animating Dynamical Systems: a Guide to XPPAUT for Researchers and Students, SIAM, 2002. |
| [53] | K. Engelborghs, T. Luzyanina, G. Samaey, DDE-BIFTOOL v. 2.00: a Matlab Package for Bifurcation Analysis of Delay Differential Equations, Technical Report TW-330, Department of Computer Science, K.U. Leuven, Leuven, Belgium, 2001. |
| [54] |
C. Morris, H. Lecar, Voltage oscillations in the barnacle giant muscle fibre, Biophysical J., 35 (1981), 193-213. doi: 10.1016/S0006-3495(81)84782-0
|
| [55] | J. Rinzel, Excitation dynamics: insights from simplified membrane models, Fed. Proc, 44 (1985), 2944-2946. |
| [56] |
D. Terman, D. Wang, Global competition and local cooperation in a network of neural oscillators, Physica D, 81 (1995), 148-176. doi: 10.1016/0167-2789(94)00205-5
|
| [57] | J. Hale, S. Verduyn Lunel, Introduction to Functional Differential Equations, Springer Verlag, New York, 1993. |
| [58] | Z. Wang, Clustering of Networks with Time Delayed All-to-all Coupling, Ph.D thesis, University of Waterloo, 2017, |
| [59] |
R. Fitzhugh, Impulses and physiological states in theoretical models of nerve membrane, Biophysical J., 1 (1961), 445-466. doi: 10.1016/S0006-3495(61)86902-6
|
| [60] |
J. Nagumo, S. Arimoto, S. Yoshizawa, An active pulse transmission line simulating nerve axon, Proceeding IRE, 50 (1962), 2061-2070. doi: 10.1109/JRPROC.1962.288235
|
| [61] | M. Golubitsky, I. Stewart, D. G. Scherffer, Singularities and Groups in Bifurcation Theory, Springer-Verlag, New York, 1988. |
| [62] |
J. Wu, Symmetric functional-differential equations and neural networks with memory, Trans. Amer. Math. Soc., 350 (1998), 4799-4838. doi: 10.1090/S0002-9947-98-02083-2
|
| [63] | J. Guckenheimer, P. Holmes, Nonlinear Oscillations, Dynamical Systems and Bifurcations of Vector Fields, Springer-Verlag, New York, 1983. |
| [64] | Y. A. Kuznetsov, Elements of Applied Bifurcation Theory, Springer Science & Business Media, 2013. |
| [65] |
J. Bélair, S. A. Campbell, Stability and bifurcations of equilibria in a multiple-delayed differential equation, SIAM J. Appl. Math., 54 (1994), 1402-1424. doi: 10.1137/S0036139993248853
|
| [66] |
Z. Kilpatrick, B. Ermentrout, Sparse gamma rhythms arising through clustering in adapting neuronal networks, PLoS Comput. Biol., 7 (2011), e1002281. doi: 10.1371/journal.pcbi.1002281
|
| [67] |
S. A. Campbell, M. Waite, Multistability in coupled Fitzhugh-Nagumo oscillators, Nonlinear Anal., 47 (2001), 1093-1104. doi: 10.1016/S0362-546X(01)00249-8
|
| [68] |
F. K. Skinner, H. Bazzazi, S. A. Campbell, Two-cell to N-cell heterogeneous, inhibitory networks: precise linking of multistable and coherent properties, J. Comput. Neurosci., 18 (2005), 343-352. doi: 10.1007/s10827-005-0331-1
|
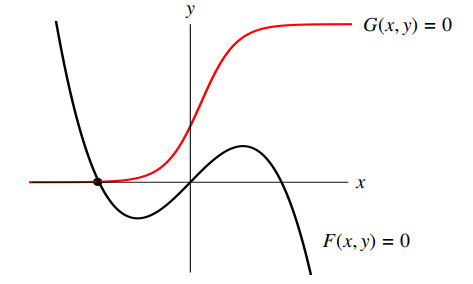
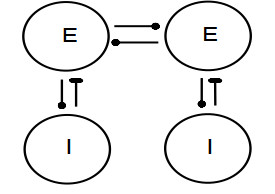
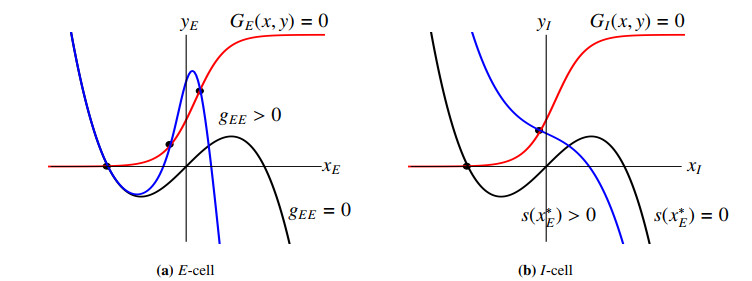
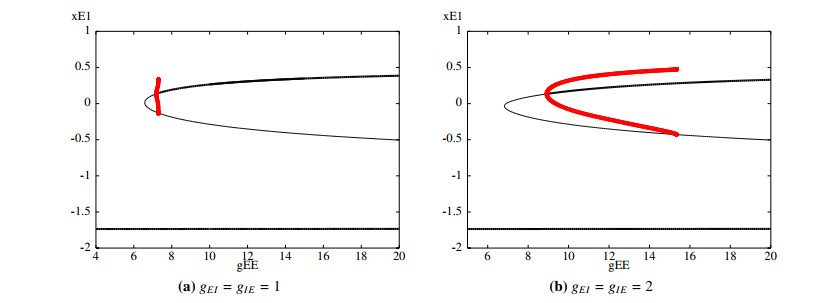
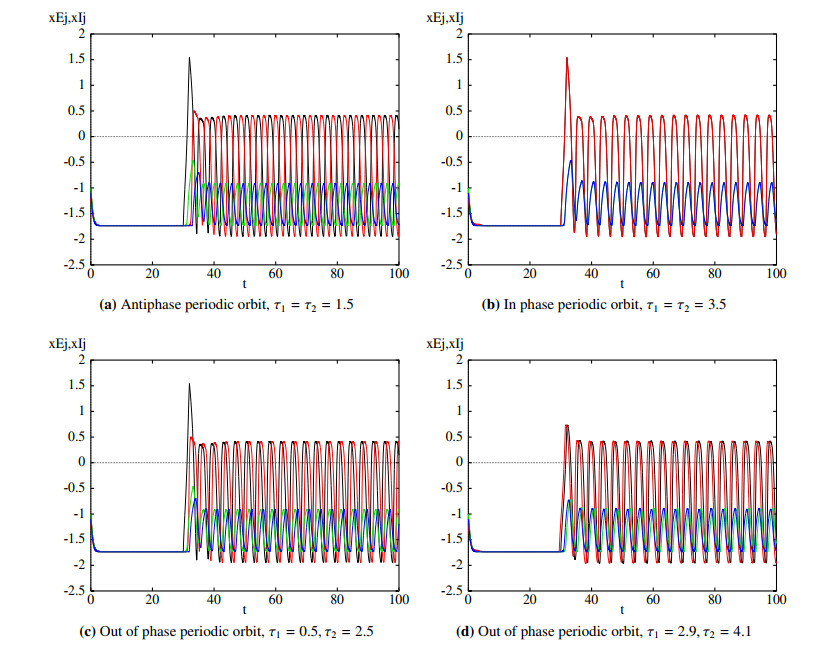
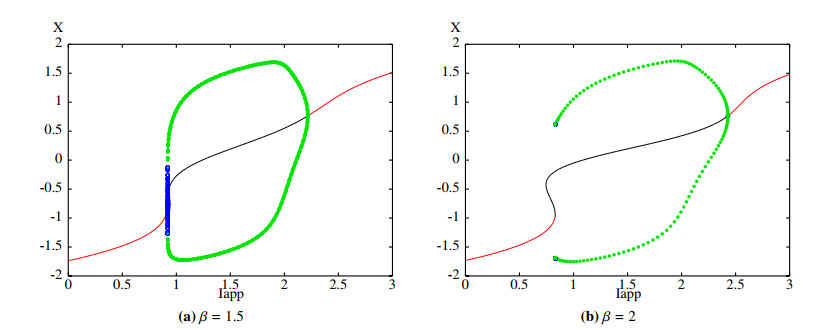
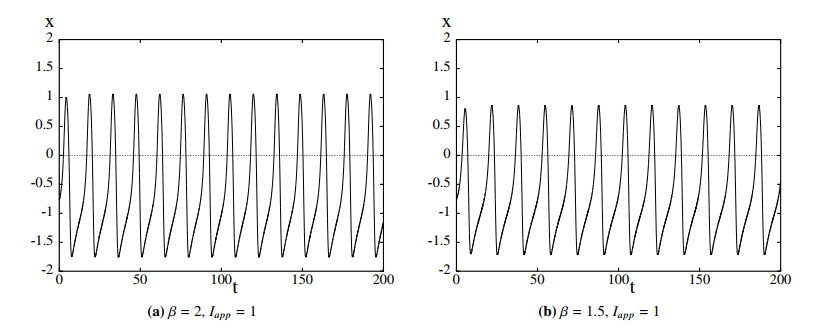
Figures(11)

Hwayeon Ryu, Sue Ann Campbell. Stability, bifurcation and phase-locking of time-delayed excitatory-inhibitory neural networks[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7931-7957. doi: 10.3934/mbe.2020403







 DownLoad:
DownLoad: