Citation: Jun Xu, Bei Wang, Zhengtao Liu, Mingchun Lai, Mangli Zhang, Shusen Zheng. miR-223-3p regulating the occurrence and development of liver cancer cells by targeting FAT1 gene[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1534-1547. doi: 10.3934/mbe.2020079

| [1] | Y. A. Ghouri, I. Mian, B. Blechacz, Cancer review: Cholangiocarcinoma, J. Carcinog., 14 (2015). |
| [2] | J. Tejeda-Maldonado, I. García-Juárez, J. Aguirre-Valadez, A. González-Aguirre, M. Vilatobá-Chapa, A. Armengol-Alonso, Diagnosis and treatment of hepatocellular carcinoma: An update, World J. Hepatol., 7 (2015), 362-376. |
| [3] | A. Kinoshita, H. Onoda, N. Fushiya, K. Koike, H. Nishino, H. Tajiri, Staging systems for hepatocellular carcinoma: Current status and future perspectives, World J. Hepatol.,7 (2015), 406-424. |
| [4] | T. Kimhofer, H. Fye, S. Taylor-Robinson, E. Holmes, Proteomic and metabonomic biomarkers for hepatocellular carcinoma: A comprehensive review, Br. J. Cancer, 112 (2015), 1141-1156. |
| [5] | J. Dunnea, A. M. Hanby, R. Poulsom, T. A. Jones, Molecular cloning and tissue expression of FAT, the human homologue of the Drosophila fat gene that is located on chromosome 4q34-q35 and encodes a putative adhesion molecule, Genomics, 30 (1995), 207-223. |
| [6] | L. Ciani, A. Patel, N. D. Allen, C. ffrench-Constant, Mice lacking the giant protocadherin mFAT1 exhibit renal slit junction abnormalities and a partially penetrant cyclopia and anophthalmia phenotype. Mol. Cell Biol., 23 (2003), 3575-3582. |
| [7] | M. Katoh, Function and cancer genomics of FAT family genes, Int. J. Oncol., 41 (2012), 1913-1918. |
| [8] | N. Kwaepila, G. Burns, A. S. Leong, Immunohistological localisation of human FAT1 (hFAT) protein in 326 breast cancers. Does this adhesion molecule have a role in pathogenesis? Pathology, 38 (2006), 125-131. |
| [9] | C. E. de Bock, A. Ardjmand, T. J. Molloy, S. M. Bone, D. Johnstone, D. M Campbell, The Fat1 cadherin is overexpressed and an independent prognostic factor for survival in paired diagnosis-relapse samples of precursor B-cell acute lymphoblastic leukemia, Leukemia, 26 (2012), 918-926. |
| [10] | B. Dikshit, K. Irshad, E. Madan, N. Aggarwal, C. Sarkar, P. S. Chandra, FAT1 acts as an upstream regulator of oncogenic and inflammatory pathways, via PDCD4, in glioma cells, Oncogene, 32 (2013), 3798-3808. |
| [11] | D. Valletta, B. Czech, T. Spruss, K. Ikenberg, P. Wild, Arndt Hartmann, Regulation and function of the atypical cadherin FAT1 in liver cancer, Carcinogenesis., 35 (2014), 1407-1415. |
| [12] | C. H. Hung, Y. C. Chiu, C. H. Chen, T. H. Hu, MicroRNAs in hepatocellular carcinoma: Carcinogenesis, progression, and therapeutic target, Biomed. Res. Int., 2014 (2014). |
| [13] | C. H. Yu, C. F. Xu, Y. M. Li, Association of MicroRNA-223 expression with hepatic ischemia/reperfusion injury in mice, Dig. Dis. Sci., 54 (2009), 2362. |
| [14] | A. Aucher, D. Rudnicka, D. M. Davis, MicroRNAs transfer from human macrophages to hepato-carcinoma cells and inhibit proliferation, J. Immunol., 191 (2013), 6250-6260. |
| [15] | J. Xu, C. Wu, X. Che, L. Wang, D. Yu, T. Zhang, Circulating microRNAs, miR-21, miR-122, and miR-223, in patients with liver cancer or chronic hepatitis, Mol. Carcinog., 50 (2), 136-142. |
| [16] | X. V. Qadir, W. Chen, C. Han, K. Song, J. Zhang, T. Wu, MiR-223 Deficiency Protects against Fas-Induced Hepatocyte Apoptosis and Liver Injury through Targeting Insulin-Like Growth Factor 1 Receptor, Am. J. Pathol., 185 (2015), 3141-3151. |
| [17] | S. Lee, S. Stewart, I. Nagtegaal, J. Luo, Y. Wu, G. Colditz, Differentially expressed genes regulating the progression of ductal carcinoma in situ to invasive breast cancer, Cancer Res., 72 (2012), 4574-4586. |
| [18] | R. Hou, L. Liu, S. Anees, S. Hiroyasu, N. E. S. Sibinga, The Fat1 cadherin integrates vascular smooth muscle cell growth and migration signals, J. Cell. Biol., 173 (2006), 417-429. |
| [19] | M. J. Moeller, A. Soofi, G. S. Braun, X. Li, C. Watzl, W. Kriz, Protocadherin FAT1 binds Ena/VASP proteins and is necessary for actin dynamics and cell polarization, EMBO. J., 23 (2004), 3769-3779. |
| [20] | C. Srivastava, K. Irshad, B. Dikshit, P. Chattopadhyay, C. Sarkarm, D. K. Gupta, FAT1 modulates EMT and stemness genes expression in hypoxic glioblastoma, Int. J. Cancer, 142 (2018), 805-812. |
| [21] | X. Hua, Y. Zhai, P. Kong, H. Cui, T. Yan, J. Yang, FAT1 prevents epithelial mesenchymal transition (EMT) via MAPK/ERK signaling pathway in esophageal squamous cell cancer, Cancer Lett., 397 (2017), 83-93. |
| [22] | B. G. Giray, G. Emekdas, S. Tezcan, M. Ulger, M. S. Serin, O. Sezgin, Profiles of serum microRNAs; miR-125b-5p and miR223-3p serve as novel biomarkers for HBV-positive liver cancer, Mol. Biol. Rep., 41 (2014), 4513-4519. |
| [23] | Z. B. Han, L. Zhong, M. J. Teng, J. W. Fan, H. M. Tang, J. Y. Wu, Identification of recurrence-related microRNAs in hepatocellular carcinoma following liver transplantation, Mol. Oncol., 6 (2012), 445-457. |
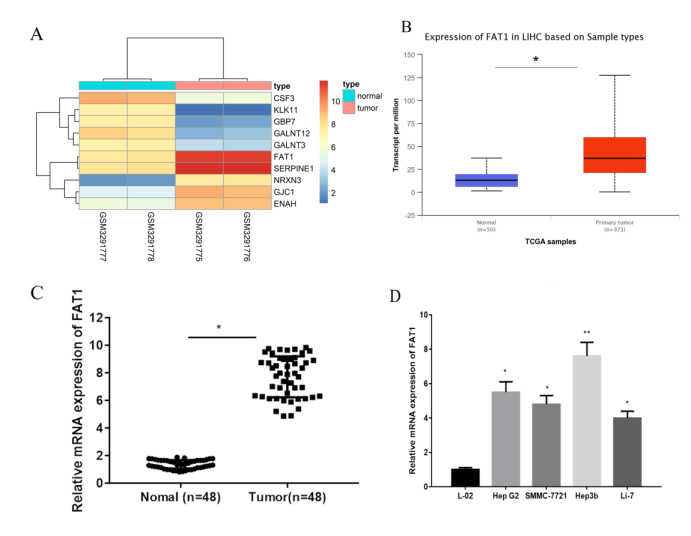
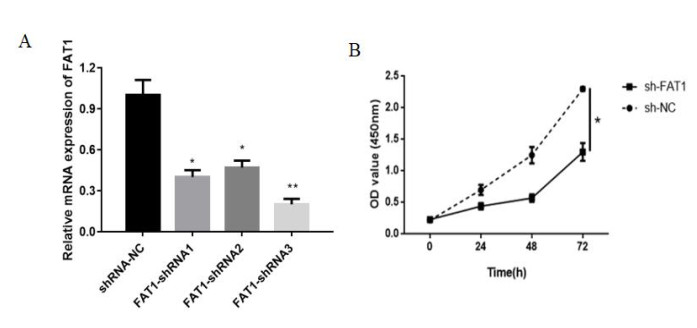
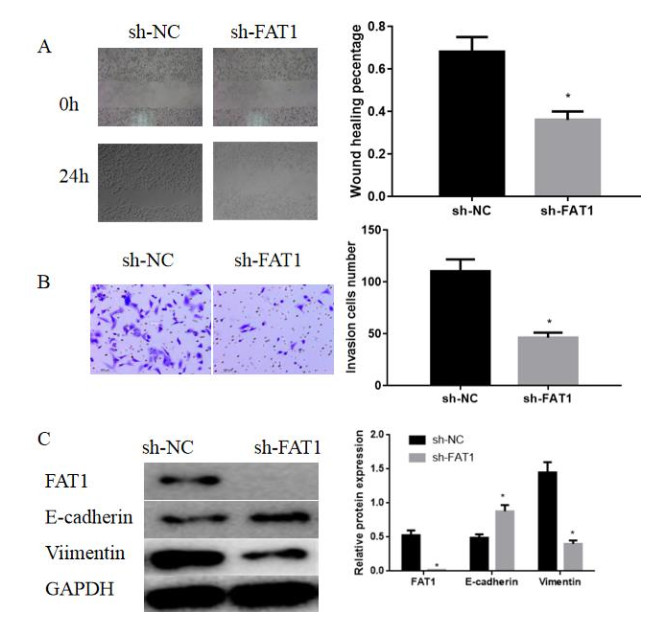

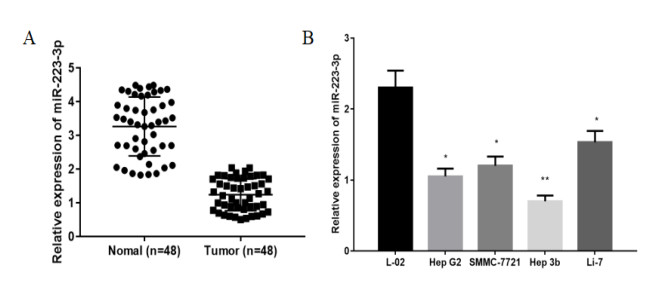
Figures(7)

Jun Xu, Bei Wang, Zhengtao Liu, Mingchun Lai, Mangli Zhang, Shusen Zheng. miR-223-3p regulating the occurrence and development of liver cancer cells by targeting FAT1 gene[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1534-1547. doi: 10.3934/mbe.2020079







 DownLoad:
DownLoad: