Citation: Jing Li, Zhen Jin, Yuan Yuan. Effect of adaptive rewiring delay in an SIS network epidemic model[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 8092-8108. doi: 10.3934/mbe.2019407

| [1] | M. Faloutsos, P. Faloutsos and C. Faloutsos, On power-law relationships of the internet topology, Comput. Commun. Rev., 29 (1999), 251-262. |
| [2] | V. Colizza, A. Barrat, M. Barthélemy, et al., The role of the airline transportation network in the prediction and predictability of global epidemics, Proc. Nati. Acad. Sci. USA, 103 (2006), 2015-2020. |
| [3] | M. E. J. Newman, The structure of scientific collaboration networks, Proc. Nati. Acad. Sci. USA, 98 (2001), 404-409. |
| [4] | M. E. J. Newman, Scientific collaboration networks. I. Network construction and fundamental results, Phys. Rev. E, 64 (2001), 016131. |
| [5] | P. L. Krapivsky, S. Redner and F. Leyvraz, Connectivity of growing random networks, Phys. Rev. Lett., 85 (2000), 4629-4632. |
| [6] | S. N. Dorogovtsev and J. F. F. Mendes, Scaling properties of scale-free evolving networks: Continuous approach, Phys. Rev. E, 63 (2001), 056125. |
| [7] | G. Kossinets and D. J. Watts, Empirical analysis of an evolving social network, Science, 311(2006), 88-90. |
| [8] | A. Clauset, C. R. Shalizi and M. E. J. Newman, Power-law distributions in empirical data, SIAM Rev., 51 (2009), 661-703. |
| [9] | M. Kivela, A. Arenäs, M. Barthélemy, et al., Multilayer network, J. Comp. Net., 2 (2014), 203-271. |
| [10] | S. Boccaletti, G. Bianconi, R. Criado, et al., The structure and dynamics of multilayer networks, Phys. Rep., 544 (2014), 1-122. |
| [11] | V. Colizza and A. Vespignani, Invasion threshold in heterogeneous metapopulation networks, Phys. Rev. Lett., 99 (2007), 148701. |
| [12] | N. Masuda, Effects of diffusion rates on epidemic spreads in metapopulation networks, New J. Phys., 12 (2010), 093009. |
| [13] | W. K. V. Chan and C. Hsu, Service scaling on hyper-networks, Serv. Science, 1 (2009), 17-31. |
| [14] | A. Trilla, G. Trilla and C. Daer, The 1918 "Spanish flu" in Spain, Clin. Infect. Dis., 47 (2008), 668-673. |
| [15] | M. A. Marra, S. J. Jones, C. R. Astell, et al., The genome sequence of the SARS-associated coronavirus, Science, 300 (2003), 1399-1404. |
| [16] |
J. S. Peiris, S. T. Lai, L. L. Poon, et al., Coronavirus as a possible cause of severe acute respiratory syndrome, Lancet, 361 (2003), 1319-1325. doi: 10.1016/S0140-6736(03)13077-2

|
| [17] | T. R. Frieden, I. Damon, B. P. Bell, et al., Ebola 2014-new challenges, new global response and responsibility, New Engl. J. Med., 371 (2014), 1177-1180. |
| [18] | W. O. Kermack and A. G. McKendrick, Contributions to the mathematical theory of epidemics, Proc. Roy. Soc. A, 115 (1927), 700-721. |
| [19] | N. T. J. Bailey, The mathematical theory of infectious diseases and its applications, Hafner Press, New York, 1975. |
| [20] | R. Anderson and R. May, Infectious disease of human: dynamic and control, Press Oxford, Oxford University, 1991. |
| [21] | M. Martcheva, Introduction to Mathematical Epidemiology, Springer-Verlag, New York, 2010. |
| [22] | I. Al-Darabsah and Y. Yuan, A time-delayed epidemic model for Ebola disease transmission, Appl. Math. Comput., 290 (2016), 307-325. |
| [23] | A. S. Klovdahl, Social networks and the spread of infectious diseases£the AIDS example, Soc. Sci. Med., 21 (1985), 1203-1216. |
| [24] | R. M. May and R. M. Andemon, Transmission dynamics of HIV infection, Nature, 326 (1987), 137-142. |
| [25] | O. Diekmann, M. C. M. De Jong and J. A. J. Metz, A deterministic epidemic model taking account of repeated contacts between the same Individuals, J. Appl. Prob., 35 (1998), 448-462. |
| [26] | Y. Moreno, R. Pastor-Satorras and A. Vespignani, Epidemic outbreaks in complex heterogeneous networks, Euro. Phys. J. B, 26 (2002), 521-529. |
| [27] | E. Volz, SIR dynamics in random networks with heterogeneous connectivity, J. Math. Biol., 56(2008) 293-310. |
| [28] | C. T. Bauch, The spread of infectious diseases in spatially structured populations: an invasory pair approximation, Math. Biosci., 198 (2005), 217-237. |
| [29] | C. Moore and M. E. J. Newman, Epidemics and percolation in small-world networks, Phys. Rev. E, 61 (2000), 5678-5682. |
| [30] | R. Pastor-Satorras and A. Vespignani, Epidemic dynamics and endemic states in complex networks, Phys. Rev. E, 63 (2001) 066117. |
| [31] | N. M. Ferguson, D. A. T. Cummings, C. Fraser, et al., Strategies for mitigating an influenza pandemic, Nature, 442 (2006), 448-452. |
| [32] | T. Gross, C. J. D. DLimaa and B. Blasius, Epidemic Dynamics on an Adaptive Network, Phys. Rev. Lett. 96 (2006), 208701. |
| [33] | D. H. Zanette and S. Risau-Gusmán, Infection spreading in a population with evolving contacts, J. Biol. Phys., 34 (2008), 135-148. |
| [34] | L. B. Shaw and I. B. Schwartz, Fluctuating epidemics on adaptive networks, Phys. Rev. E 77(2008), 066101. |
| [35] | A. Szabó-Solticzky, L. Berthouze, I. Z. Kiss, et al., Oscillating epidemics in a dynamic network model: stochastic and mean-field analysis, J. Math. Biol. 72 (2016), 1153-1176. |
| [36] | J. Li, Z. Jin, Y. Yuan, et al., A non-Markovian SIR network model with fixed infectious period and preventive rewiring, Comput. Math. Appl., 75 (2018), 3884-3902. |
| [37] | I. Z. Kiss, L. Berthouze, T. J. Taylor, et al., Modelling approaches for simple dynamic networks and applications to disease transmission models, Proc. R. Soc. A, 468 (2012), 1332-1355. |
| [38] | T. Rogers, W. Clifford-Brown, C. Mills, et al., Stochastic oscillations of adaptive networks: application to epidemic modelling, J. Stat. Mech., 2012 (2012), P08018. |
| [39] | S. Risau-Gusman and D. H. Zanette, Contact switching as a control strategy for epidemic outbreaks, J. Theor. Biol., 257 (2009), 52-60. |
| [40] | M. J. Keeling, The effects of local spatial structure on epidemiological invasions, Proc. R. Soc. Lond. B, 266 (1999), 859-867. |
| [41] | X. Zhang, C. Shan, Z. Jin, et al., Complex dynamics of epidemic models on adaptive networks,Journal of differential equations, J. Differ. Equations, 266 (2019), 803-832. |
| [42] | J. Graef, M. Li and L. Wang, A study on the effects of disease caused death in a simple epidemic model, in Dyn. Syst. Differ. Equations (eds. W. Chen and S Hu), Southwest Missouri State University Press, (1998), 288-300. |
| [43] | M. Y. Li, W. Liu, C. Shan, et al., Turning Points And Relaxation Oscillation Cycles in Simple Epidemic Models, SIAM J. Appl. Math., 76 (2016), 663-687. |
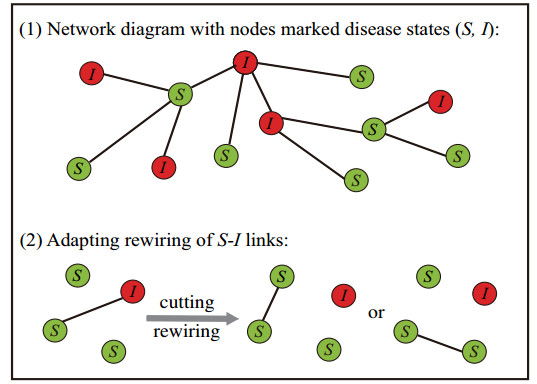
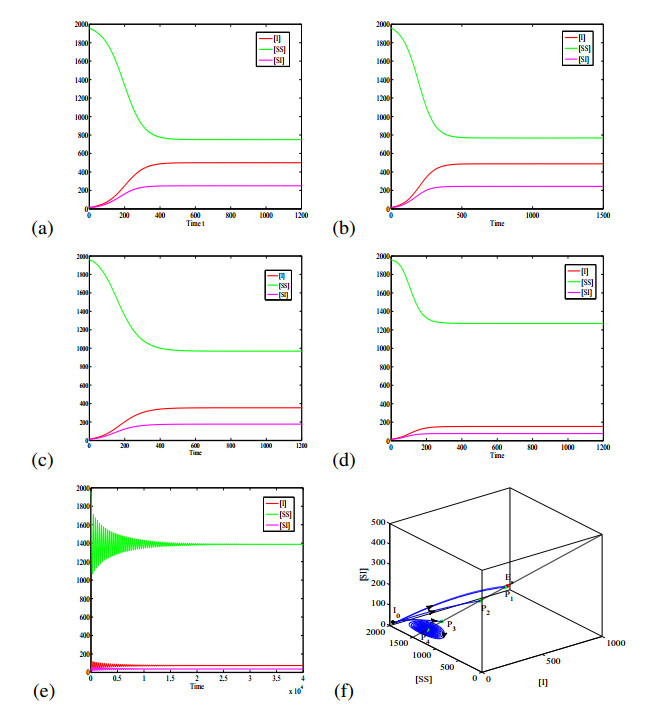
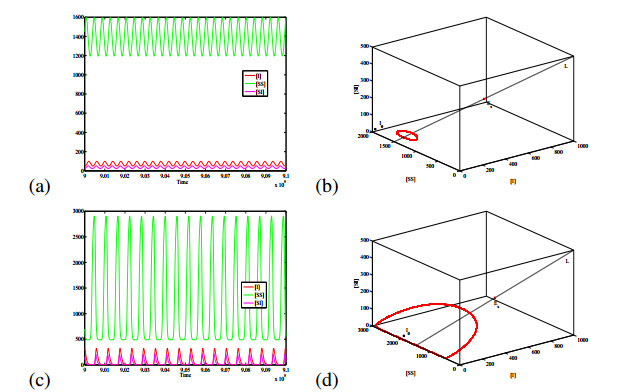
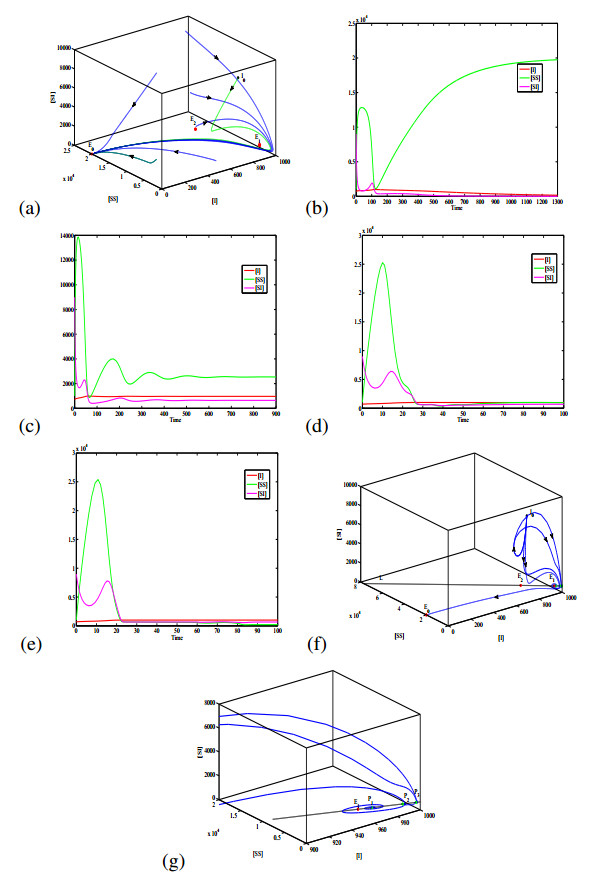
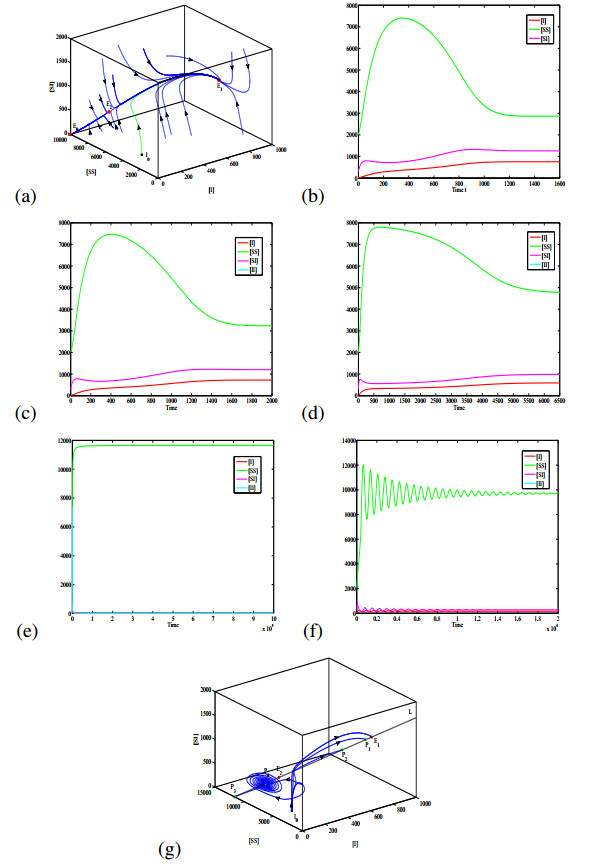
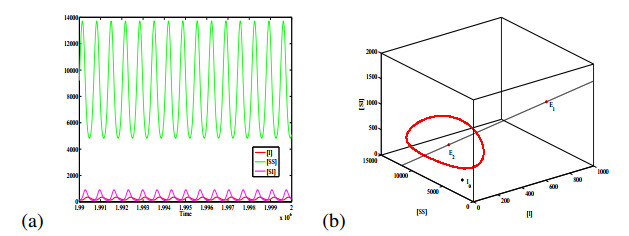
Figures(6) / Tables(1)

Jing Li, Zhen Jin, Yuan Yuan. Effect of adaptive rewiring delay in an SIS network epidemic model[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 8092-8108. doi: 10.3934/mbe.2019407







 DownLoad:
DownLoad: