An SIS epidemic model with time delay and stochastic perturbation on scale-free networks is established in this paper. And we derive sufficient conditions guaranteeing extinction and persistence of epidemics, respectively, which are related to the basic reproduction number $ R_0 $ of the corresponding deterministic model. When $ R_0 < 1 $, almost surely exponential extinction and $ p $-th moment exponential extinction of epidemics are proved by Razumikhin-Mao Theorem. Whereas, when $ R_0 > 1 $, the system is persistent in the mean under sufficiently weak noise intensities, which indicates that the disease will prevail. Finally, the main results are demonstrated by numerical simulations.
Citation: Meici Sun, Qiming Liu. An SIS epidemic model with time delay and stochastic perturbation on heterogeneous networks[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 6790-6805. doi: 10.3934/mbe.2021337
An SIS epidemic model with time delay and stochastic perturbation on scale-free networks is established in this paper. And we derive sufficient conditions guaranteeing extinction and persistence of epidemics, respectively, which are related to the basic reproduction number $ R_0 $ of the corresponding deterministic model. When $ R_0 < 1 $, almost surely exponential extinction and $ p $-th moment exponential extinction of epidemics are proved by Razumikhin-Mao Theorem. Whereas, when $ R_0 > 1 $, the system is persistent in the mean under sufficiently weak noise intensities, which indicates that the disease will prevail. Finally, the main results are demonstrated by numerical simulations.

| [1] |
A. L. Barabási, R. Alber, Emergence of scaling in random networks, Science, 286 (1999), 509–512. doi: 10.1126/science.286.5439.509

|
| [2] | M. Barthelemy, A. Barrat, R. Pastor-Satorras, A. Vespignani, Dynamical patterns of epidemic outbreaks in complex heterogeneous networks, J. Theor. Biol., 235 (2004), 275–288. |
| [3] |
T. Zhou, J. Liu, W. Bai, G. Chen, B. wang, Behaviors of susceptible-infected epidemics on scale-free networks with identical infectivity, Phys. Rev. E, 74 (2006), 056109. doi: 10.1103/PhysRevE.74.056109
|
| [4] |
R. Pastor-Satorras, A. Vespignani, Epidemic spreading in scale-free networks, Phys. Rev. Lett., 86 (2001), 3200–3203. doi: 10.1103/PhysRevLett.86.3200
|
| [5] |
R. Pastor-Satorras, A. Vespignani, Epidemic dynamics and endemic states in complex networks, Phys. Rev. E, 63 (2001), 066117. doi: 10.1103/PhysRevE.63.066117
|
| [6] | Y. Moreno, R. Pastor-Satorras, A. Vespignanil, Epidemic outbreaks in complex heterogeneous networks, Eur. Phys. J. B, 26 (2002), 521–529. |
| [7] |
K. Li, G. Zhu, Z. Ma, L. Chen, Dynamic stability of an SIQS epidemic network and its optimal control, Commun. Nonlinear Sci. Numer. Simul., 66 (2019), 84–95. doi: 10.1016/j.cnsns.2018.06.020
|
| [8] |
J. Liu, T. Zhang, Epidemic spreading of an SEIRS model in scale-free networks, Commun. Nonlinear Sci. Numer. Simul., 16 (2011), 3375–3384. doi: 10.1016/j.cnsns.2010.11.019
|
| [9] |
T. Li, Y. Wang, Z. Guan, Spreading dynamics of a SIQRS epidemic model on scale-free networks, Commun. Nonlinear Sci. Numer. Simul., 19 (2014), 686–692. doi: 10.1016/j.cnsns.2013.07.010
|
| [10] |
L. Wang, G. Dai, Global stability of virus spreading in complex heterogeneous networks, SIAM J. Appl. Math., 68 (2008), 1495–1502. doi: 10.1137/070694582
|
| [11] |
A. d'Onofrio, A note on global behavior of the network-based SIS epidemic model, Nonlinear Anal.: Real World Appl., 9 (2008), 1567–1572. doi: 10.1016/j.nonrwa.2007.04.001
|
| [12] |
Q. Liu, C. Deng, M. Sun, The analysis of an epidemic model with time delay on scale-free networks, Physica A, 410 (2014), 79–87. doi: 10.1016/j.physa.2014.05.010
|
| [13] |
Q. Liu, M. Sun, Dynamics of epidemic spreading model with distributed delay on heterogeneous network, J. Biol. Syst., 25 (2017), 173–183. doi: 10.1142/S0218339017500097
|
| [14] | H. Kang, Y. Lou, G. Chen, S. Chu, X. Fu, Epidemic spreading and global stability of a new SIS model with delay on heterogenous network, J. Biol. Syst., 23 (2015), 577–595. |
| [15] |
H. Kang, X. Fu, Epidemic spreading and global stability of an SIS model with an infective vector on complex networks, Commun. Nonlinear Sci. Numer. Simul., 27 (2015), 30–39. doi: 10.1016/j.cnsns.2015.02.018
|
| [16] |
S. Bonaccorsi, S. Ottaviano, Epidemics on networks with heterogeneous population and stochastic infection rates, Math. Biosci., 279 (2016), 43–52. doi: 10.1016/j.mbs.2016.07.002
|
| [17] |
A. Krause, L. Kurowski, K. Yawar, R. A. Van Gorder, Stochastic epidemic metapopulation models on networks: sis dynamics and control strategies, J. Theor. Biol., 449 (2018), 35–52. doi: 10.1016/j.jtbi.2018.04.023
|
| [18] | H. Yang, Z. Jin, Stochastic SIS epidemic model on network with Lévy noise, Stoch. Anal. Appl., 2021, DOI: 10.1080/07362994.2021.1930051. |
| [19] |
C. Li, C. Tsai, S. Yang, Analysis of epidemic spreading of an SIRS model in complex heterogeneous networks, Commun. Nonlinear Sci. Numer. Simul., 19 (2014), 1042–1054. doi: 10.1016/j.cnsns.2013.08.033
|
| [20] |
G. Zhu, X. Fu, G. Chen, Global attractivity of a network-based epidemics SIS model with nonlinear infectivity, Commun. Nonlinear Sci. Numer. Simul., 17 (2012), 2588–2594. doi: 10.1016/j.cnsns.2011.08.039
|
| [21] |
R. Olinky, L. Olinky, Unexpected epidemic threshold in heterogeneous networks: the role of disease transmission, Phys. Rev. E, 70 (2004), 030902. doi: 10.1103/PhysRevE.70.030902
|
| [22] |
Q. Luo, X. Mao, Stochastic population dynamics under regime switching, J. Math. Anal. Appl., 334 (2007), 69–84. doi: 10.1016/j.jmaa.2006.12.032
|
| [23] |
A. Gray, D. Greenhalgh, L. Hu, X. Mao, J. Pan, A stochastic differential equation SIS epidemic model, SIAM J. Appl. Math., 71 (2011), 876–902. doi: 10.1137/10081856X
|
| [24] | L. Arnold, Stochastic Differential Equations: Theory and Applications, Wiley, New York, 1974. |
| [25] | X. Mao, Stochastic Differential Equations and Applications, 2$^{nd}$ edition, Woodhead Publishing, Cambridge, 2007. |
| [26] |
C. Ji, D. Jiang, N. Shi, Multigroup SIR epidemic model with stochastic perturbation, Physica A, 390 (2011), 1747–1762. doi: 10.1016/j.physa.2010.12.042
|
| [27] |
H. Zhang, X. Fu, Spreading of epidemics on scale-free networks with nonlinear infectivity, Nonlinear Anal., 70 (2009), 3273–3278. doi: 10.1016/j.na.2008.04.031
|
| [28] |
D. J. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001), 525–546. doi: 10.1137/S0036144500378302
|
| [29] | J. R. C. Piqueira, V. O. Araujo, A modified epidemiological model for computer viruses, Appl. Math. Comput., 213 (2009), 355–360. |
| [30] |
J. R. C. Piqueira, M. A. M. Cabrera, C. M. Batistela, Malware propagation in clustered computer networks, Physica A, 573 (2021), 125958. doi: 10.1016/j.physa.2021.125958
|
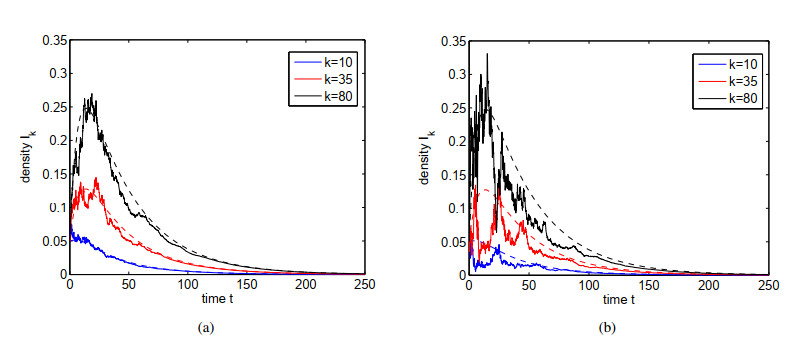
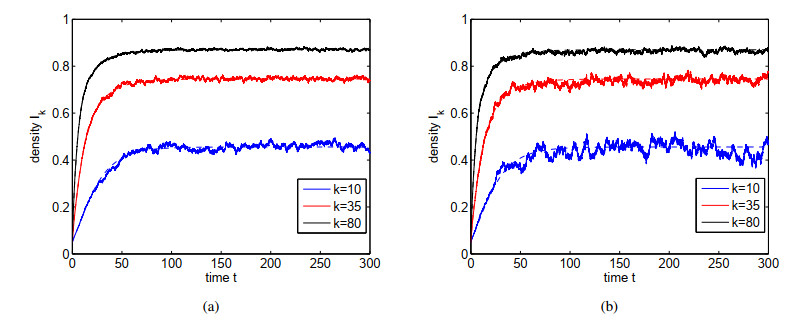
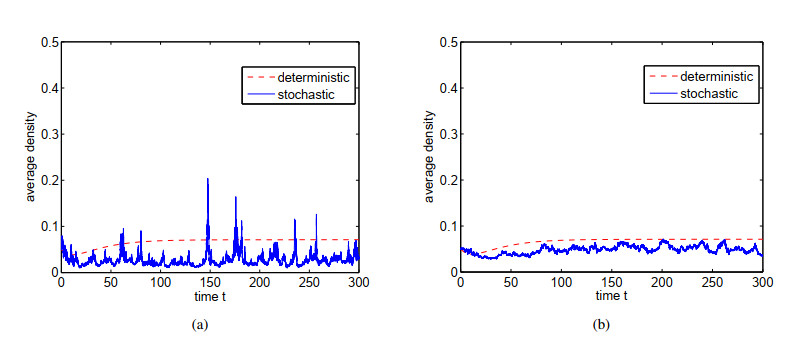
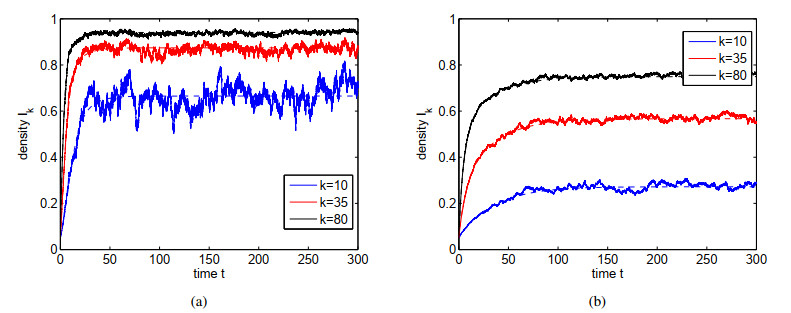
Figures(4)

Meici Sun, Qiming Liu. An SIS epidemic model with time delay and stochastic perturbation on heterogeneous networks[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 6790-6805. doi: 10.3934/mbe.2021337







 DownLoad:
DownLoad: