Citation: Ahmed Nagah, Asmaa Amer, Xinan Zhang. The effect of cigarette smoking on lung cancer evolution[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7771-7788. doi: 10.3934/mbe.2019390

| [1] | D. M. Parkin, F. Bray, J. Ferlay, et al., Global cancer statistics, 2002 , CA Cancer J. Clin., 55 (2005), 74–108. |
| [2] | R. L. Siegel, K. D. Miller and A. Jemal, Cancer statistics, 2015, CA Cancer J. Clin., 65 (2015), 5–29. |
| [3] | M. Malvezzi, G. Carioli, P. Bertuccio, et al., European cancer mortality predictions for the year 2016 with focus on leukaemias, Ann. Oncol., 27 (2016), 725–731. |
| [4] | J. Ferlay, I. Soerjomataram, Dikshit, et al., Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012, Int. J. Cancer, 136 (2015), E359–E386. |
| [5] | J. Malhotra, M. Malvezzi, E. Negri, et al., Risk factors for lung cancer worldwide, Eur. Resp. J., 48 (2016), 889–902. |
| [6] | S. Sun, J. H. Schiller and A. F. Gazdar, Lung cancer in never smokersa different disease, Nat. Rev. Cancer, 7 (2007), 778–790. |
| [7] | A. Blair and L. B. Freeman, Lung cancer among nonsmokers, Epidemiology, 17 (2006), 601–613. |
| [8] | R. W. Field and B. L. Withers, Occupational and environmental causes of lung cancer, Clin. Chest Med., 33 (2012), 681–703. |
| [9] | C. S. D. Cruz, L. T. Tanoue and R. A. Matthay, Lung cancer: Epidemiology, etiology, and prevention, Clin. Chest Med., 32 (2011), 605–644. |
| [10] | S. Wu, S. Powers, W. Zhu, et al., Substantial contribution of extrinsic risk factors to cancer development, Nature, 529 (2016), 43–47. |
| [11] | G. Ray, D. E. Henson and A. M. Schwartz, Cigarette smoking as a cause of cancers other than lung cancer: An exploratory study using the Surveillance, Epidemiology, and End Results Program, Chest, 138 (2010), 491–499. |
| [12] | S. D. Coello, A. C. De Len, F. B. Ojeda, et al., High density lipoprotein cholesterol increases with living altitude, Int. J. Epidemiol., 29 (2000), 65–70. |
| [13] | V. L. Roger, M. E. Farkouh, S. A. Weston, et al., Sex differences in evaluation and outcome of unstable angina, JAMA, 283 (2000), 646–652. |
| [14] | M. Mori, M. Hara, I. Wada, et al., Prospective study of hepatitis B and C viral infections, cigarette smoking, alcohol consumption, and other factors associated with hepatocellular carcinoma risk in Japan, Am. J. Epidemiol., 151 (2000), 131–139. |
| [15] | K. Husgafvel-Pursiainen, P. Boffetta, A. Kannio, et al., p53 mutations and exposure to environ-mental tobacco smoke in a multicenter study on lung cancer, Cancer Res., 60 (2000), 2906–2911. |
| [16] | T. L. Holmen, E. Barrett-Connor, J. Holmen, et al., Health problems in teenage daily smokers versus nonsmokers, Norway, 1995–1997: The Nord-Trndelag Health Study, Am. J. Epidemiol., 151 (2000), 148–155. |
| [17] | G. Launoy, C. Milan, J. Faivre, et al., Tobacco type and risk of squamous cell cancer of the oesophagus in males: A French multicentre case-control study, Int. J. Epidemiol., 29 (2000), 36–42. |
| [18] | F. Barbone, M. Bovenzi, F. Cavallieri, et al., Cigarette smoking and histologic type of lung cancer in men, Chest, 112 (1997), 1474–1479. |
| [19] | L. M. Nelson, V. McGuire, W. T. Longstreth Jr, et al., Population-based case-control study of amyotrophic lateral sclerosis in western Washington State. I. Cigarette smoking and alcohol consumption, Am. J. Epidemiol., 151 (2000), 156–163. |
| [20] | K. Leffondr, M. Abrahamowicz, J. Siemiatycki, et al., Modeling smoking history: A comparison of different approaches, Am. J. Epidemiol., 156 (2002), 813–823. |
| [21] | R. Doll and R. Peto, Cigarette smoking and bronchial carcinoma: dose and time relationships among regular smokers and lifelong non-smokers, J. Epidemiol. Commun. Health, 32 (1978), 303–313. |
| [22] | W. D. Flanders, C. A. Lally, B. P. Zhu, et al., Lung cancer mortality in relation to age, duration of smoking, and daily cigarette consumption: results from Cancer Prevention Study II, Cancer Res., 63 (2003), 6556–6562. |
| [23] | C. O. Nordling, A new theory on the cancer-inducing mechanism, British J. Cancer, 7 (1953), 68–72. |
| [24] | P. Armitage and R. Doll, The age distribution of cancer and a multi-stage theory of carcinogenesis, British J. Cancer, 8 (1954), 1–12. |
| [25] | I. Tomlinson and W. Bodmer, Selection, the mutation rate and cancer: ensuring that the tail does not wag the dog, Nat. Med., 5 (1999), 11–12. |
| [26] | K. Anderson, C. Lutz, F. W. Van Delft, et al., Genetic variegation of clonal architecture and propagating cells in leukaemia, Nature, 469 (2011), 356–361. |
| [27] | M. Gerlinger, S. Horswell, J. Larkin, et al., Genomic architecture and evolution of clear cell renal cell carcinomas defined by multiregion sequencing, Nat. Genet., 46 (2014), 225–233. |
| [28] | M. R. Stratton, P. J. Campbell and P. A. Futreal, The cancer genome, Nature, 458 (2009), 719–724. |
| [29] | L. D. Wood, D. W. Parsons, S. Jones, et al., The genomic landscapes of human breast and colorectal cancers, Science, 318 (2007), 1108–1113. |
| [30] | H. M. Byrne, Dissecting cancer through mathematics: from the cell to the animal model, Nat. Rev. Cancer, 10 (2010), 221–230. |
| [31] | S. H. Moolgavkar, A. Dewanji and G. Luebeck, Cigarette smoking and lung cancer: reanalysis of the British doctor's data, J. Nat. Cancer Inst., 81 (1989), 415–420. |
| [32] | A. S. Whittemore, Effect of cigarette smoking in epidemiological studies of lung cancer, Statist. Med., 7 (1988), 223–238. |
| [33] | M. Gaffney and B. Altshuler, Examination of the Role of Cigarette Smoke in Lung Carcinogenesis Using Multistage Models, J. Nat. Cancer Inst., 80 (1988), 925–931. |
| [34] | S. H. Moolgavkar and A. G. Knudson, Mutation and cancer: A model for human carcinogenesis, J. Nat. Cancer Inst., 66 (1981), 1037–1052. |
| [35] | R. Meza, W. D. Hazelton, G. A. Colditz, et al., Analysis of lung cancer incidence in the nurses health and the health professionals follow-up studies using a multistage carcinogenesis model, Cancer Causes Control, 19 (2008), 317–328. |
| [36] | Surveillance, Epidemiology, and End Results (SEER) Program (www.seer.cancer.gov) SEER*Stat Database: Incidence-SEER 13 Regs Research Data, Nov 2012 Sub (1992-2010) $<$Katrina/Rita Population Adjustment$>$, National Cancer Institute, DCCPS, Surveillance Research Program, Surveillance Systems Branch, released April 2013, based on the November 2012 submission. |
| [37] | D. R. Lewis, D. P. Check, N. E. Caporaso, et al., US lung cancer trends by histologic type, Cancer, 120 (2014), 2883–2892. |
| [38] | C. Bain, D. Feskanich, F. E. Speizer, et al., Lung cancer rates in men and women with comparable histories of smoking, J. Nat. Cancer Inst., 96 (2004), 826–834. |
| [39] | D. Hanahan and R. A. Weinberg, Hallmarks of cancer: The next generation, Cell, 144 (2011), 646–674. |
| [40] | T. E. Harris, The theory of branching processes, Prentice-Hall Applied Mathematics Series, Englewood CLiffs, NJ, 2002. |
| [41] | C. Tomasetti, L. Marchionni, M. A. Nowak, et al., Only three driver gene mutations are required for the development of lung and colorectal cancers, Proceed. Nat. Aca. Sci., 112 (2015), 118–123. |
| [42] | L. Li, T. Tian and X. Zhang, The impact of radiation on the development of lung cancer, J. Theor. Biol., 428 (2017), 147–152. |
| [43] | L. A. Loeb, K. R. Loeb and J. P. Anderson, Multiple mutations and cancer, Proceed. Nat. Aca. Sci., 100 (2003), 776–781. |
| [44] | L. A. Loeb, Mutator phenotype may be required for multistage carcinogenesis, Cancer Res., 51 (1991), 3075–3079.45. H. Schllnberger, N. Beerenwinkel, R. Hoogenveen, et al., Cell selection as driving force in lung and colon carcinogenesis, Cancer Res., 70 (2010), 6797–6803. |
| [45] | 46. W. F. Heidenreich, E. G. Luebeck and S. H. Moolgavkar, Some properties of the hazard function of the twomutation clonal expansion model, Risk Anal., 17 (1997), 391–399. |
| [46] | 47. R. Meza, J. Jeon, S. H. Moolgavkar, et al., Age-specific incidence of cancer: Phases, transitions, and biological implications, Proceed. Nat. Aca. Sci., 105 (2008), 16284–16289. |
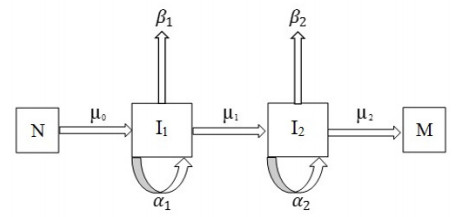
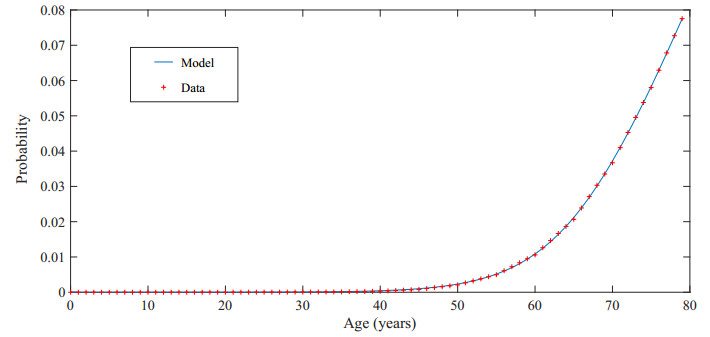
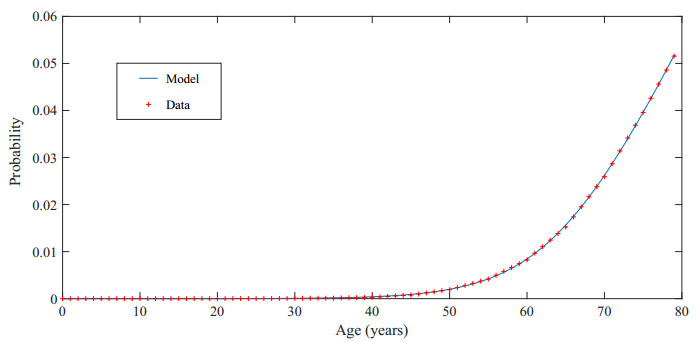
Figures(3) / Tables(10)

Ahmed Nagah, Asmaa Amer, Xinan Zhang. The effect of cigarette smoking on lung cancer evolution[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7771-7788. doi: 10.3934/mbe.2019390







 DownLoad:
DownLoad: