Citation: Giulia Pozzi, Stefano Marchesi, Giorgio Scita, Davide Ambrosi, Pasquale Ciarletta. Mechano-biological model of glioblastoma cells in response to osmotic stress[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2795-2810. doi: 10.3934/mbe.2019139

| [1] | M. Goodenberger and R. Jenkins, Genetics of adult glioma, Cancer Genet., 205 (2012), 613–621. |
| [2] | H. Ohgaki and P. Kleihues, The definition of primary and secondary glioblastoma, Clin. Cancer Res., pages clincanres–3002, 2012. |
| [3] | D. Orringer, D. Lau, S. Khatri, et al., Extent of resection in patients with glioblastoma: limiting factors, perception of resectability, and effect on survival, J. Neurosurg., 117 (2012), 851–859. |
| [4] | B. Mukherjee, B. McEllin, C. V. Camacho, et al., Egfrviii and dna double-strand break repair: a molecular mechanism for radioresistance in glioblastoma. Cancer Res., 69 (2009), 4252–4259. |
| [5] | S. Carlsson, S. Brothers and C. Wahlestedt. Emerging treatment strategies for glioblastoma multiforme. EMBO Mol. Med., e201302627, 2014. |
| [6] | M. C. Mabray, R. F. Barajas and S. Cha, Modern brain tumor imaging, Brain Tumor Res. Treat., 3 (2015), 8–23. |
| [7] | J. Folkman and M. Hochberg, Self-regulation of growth in three dimensions, J. Exp. Med., 138 (1973), 745–753. |
| [8] | R. M. Sutherland, Cell and environment interactions in tumor microregions: the multicell spheroid model, Science, 240 (1988), 177–184. |
| [9] | R. M. Sutherland, J. A. McCredie and W. R. Inch, Growth of multicell spheroids in tissue culture as a model of nodular carcinomas, J. Natl. Cancer Inst., 46 (1971), 113–120. |
| [10] | G. Helmlinger, P. Netti, H. Lichtenbeld, et al., Solid stress inhibits the growth of multicellular tumor spheroids, Nature Biotech., 15 (1997), 778. |
| [11] | G. Cheng, J. Tse, R. Jain, et al., Micro-environmental mechanical stress controls tumor spheroid size and morphology by suppressing proliferation and inducing apoptosis in cancer cells, PLoS One, 4 (2009), e4632. |
| [12] | F. Montel, M. Delarue, J. Elgeti, et al., Stress clamp experiments on multicellular tumor spheroids. Phys. Rev. Lett., 107 (2011), 188102. |
| [13] | F. Montel, M. Delarue, J. Elgeti, et al., Isotropic stress reduces cell proliferation in tumor spheroids. New J. Phys., 14 (2012), 055008. |
| [14] | P. Mascheroni, C. Stigliano, M. Carfagna, et al., Predicting the growth of glioblastoma multiforme spheroids using a multiphase porous media model, Biomech. Model. Mechanobiol., 15 (2016), 1215–1228. |
| [15] | B. Bober, J. Love, S. Horton, et al., Actin–myosin network influences morphological response of neuronal cells to altered osmolarity. Cytoskeleton, 72 (2015), 193–206. |
| [16] | F. Guilak, G. Erickson and H. Ting-Beall, The effects of osmotic stress on the viscoelastic and physical properties of articular chondrocytes, Biophys. J., 82 (2002), 720–727. |
| [17] | C. La Porta, A. Ghilardi, M. Pasini, et al., Osmotic stress affects functional properties of human melanoma cell lines, Eur. Phys. J. Plus, 130 (2015), 64. |
| [18] | A. Taloni, A. A. Alemi, E. Ciusani, et al., Mechanical properties of growing melanocytic nevi and the progression to melanoma. PloS One, 9 (2014), e94229. |
| [19] | G. Tao, L. Rott, A. Lowe, et al., Hyposmotic stress induces cell growth arrest via proteasome activation and cyclin/cyclin-dependent kinase degradation, J. Biol. Chem., 277 (2002), 19295– 19303. |
| [20] | M. Delarue, F. Montel, D. Vignjevic, et al., Compressive stress inhibits proliferation in tumor spheroids through a volume limitation, Biophys. J., 107 (2014), 1821–1828. |
| [21] | K. Tsujita, T. Takenawa and T. Itoh, Feedback regulation between plasma membrane tension and membrane-bending proteins organizes cell polarity during leading edge formation, Nature Cell Biol., 17 (2015), 749. |
| [22] | J. Schindelin, I. Arganda-Carreras, E. Frise, et al., Fiji: an open-source platform for biologicalimage analysis, Nat. Methods, 9 (2012), 676. |
| [23] | G. Schmid-Schönbein, K. Sung, H. Tözeren, et al., Passive mechanical properties of human leukocytes, Biophys. J., 36 (1981), 243–256. |
| [24] | D. Theret, M. Levesque, M. Sato, et al., The application of a homogeneous half-space model in the analysis of endothelial cell micropipette measurements, J. Biomechan. Eng., 110 (1988), 190–199. |
| [25] | R. W. Ogden, Non-linear elastic deformations. Courier Corporation, 1997. |
| [26] | T. C. Gasser, R. W. Ogden and G. A. Holzapfel, Hyperelastic modelling of arterial layers with distributed collagen fibre orientations, J. R. Soc. Interface, 3 (2005), 15–35. |
| [27] | A. Agosti, D. Ambrosi and S. Turzi, Strain energy storage and dissipation rate in active cell mechanics, Phys. Rev. E, 97 (2018), 052410. |
| [28] | R. Rivlin and J. Ericksen, Stress-deformation relations for isotropic materials, Arch. Ration. Mech. Anal., 4 (1955), 323–425. |
| [29] | A. Spencer, Part iii. theory of invariants, Continuum Physics, 1 (1971), 239–353. |
| [30] | D. Ambrosi, S. Pezzuto, D. Riccobelli, et al., Solid tumors are poroelastic solids with a chemomechanical feedback on growth, J. Elasticity, 129 (2017), 107–124. |
| [31] | A. V. Melnik, X. Luo and R. W. Ogden, A generalised structure tensor model for the mixed invariant i8, Int. J. Non-Lin. Mech., 107 (2018), 137–148. |
| [32] | C. Lim, E. Zhou and S. Quek, Mechanical models for living cells-a review, J. Biomech., 39 (2006), 195–216. |
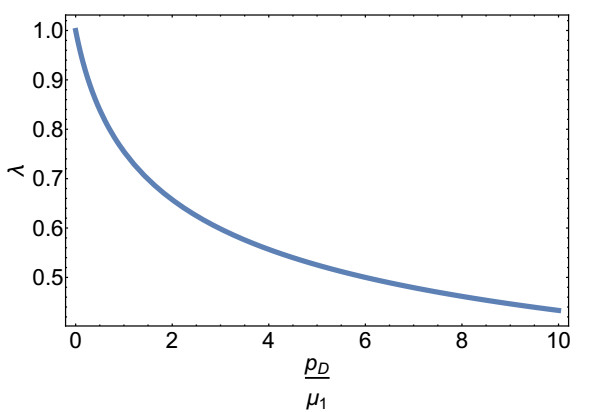
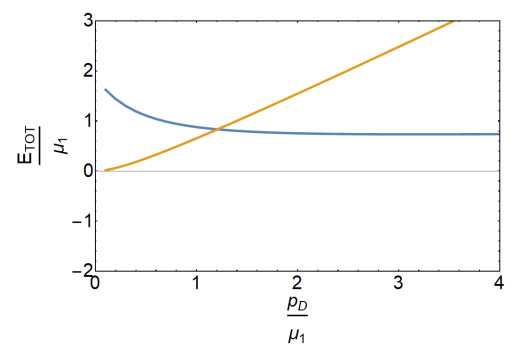
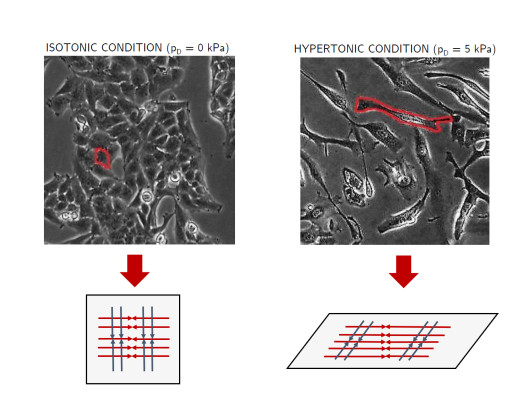
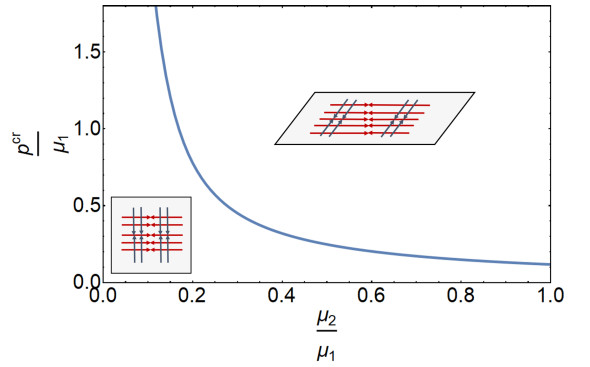
Figures(8)

Giulia Pozzi, Stefano Marchesi, Giorgio Scita, Davide Ambrosi, Pasquale Ciarletta. Mechano-biological model of glioblastoma cells in response to osmotic stress[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2795-2810. doi: 10.3934/mbe.2019139







 DownLoad:
DownLoad: