Citation: Li-zhen Du, Shanfu Ke, Zhen Wang, Jing Tao, Lianqing Yu, Hongjun Li. Research on multi-load AGV path planning of weaving workshop based on time priority[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2277-2292. doi: 10.3934/mbe.2019113

| [1] | H. Fazlollahtabar and M. Saidi-Mehrabad, Models for AGVs' scheduling and routing, Autonomous Guided Vehicles, 20 (2015), 1–15. |
| [2] | G. M. Li, X. Y. Li, L. Gao, et al., Tasks assigning and sequencing of multiple AGVs based on an improved harmony search algorithm, J. Amb. Inter. Hum. Comp., 2018. Available from: DOI, 10.1007/s12652-018-1137-0. |
| [3] | D. H. Lee, Resource-based task allocation for multi-robot systems, Robot Auton. Syst., 103 (2018), 151–161. |
| [4] | T. J. Chen, Y. Sun, W. Dai, et al., On the shortest and conflict-free path planning of multi-AGV system based on dijkstra algorithm and the dynamic time-window method, Adv. Mater. Res., 645 (2013), 267–271. |
| [5] | A. Kaplan, N. Kingry, P. Uhing, et al., Time-optimal path planning with power schedules for a solar-powered ground robot, IEEE T. Autom. Sci. Eng., 14 (2017), 1235–1244. |
| [6] | N. Smolic-Rocak, S. Bogdan, Z. Kovacic, et al., Time windows based dynamic routing in multi-AGV systems, IEEE T. Autom. Sci. Eng., 7 (2010), 151–155. |
| [7] | K. G. Huo, Y. Q. Zhang, Z. H. Hu, et al., Research on scheduling problem of multi-load AGV at automated container terminal, J. Dalian Univ. Techno., 6 (2016), 24–251. |
| [8] | G. M. Li, B. Zeng, W. Liao, et al., A new AGV scheduling algorithm based on harmony search for material transfer in a real-world manufacturing system, Adv. Mech. Eng., 10 (2018), 1–13. |
| [9] | M. Mousavi, H. J. Yap, S. N. Musa, et al., Multi-objective AGV scheduling in an FMS using a hybrid of genetic algorithm and particle swarm optimization, PLoS One, 12 (2017), 1–24. |
| [10] | F. Hamed and M. Nezam, An optimal path in a bi-criteria AGV-based flexible job shop manufacturing system having uncertain parameters, Int. J. Ind. Syst. Eng., 13 (2010), 27–55. |
| [11] | H. Fazlollahtabara and M. Saidi-Mehrabad, Optima path in an intelligent AGV based manufacturing system, Transp. Lett., 7 (2015), 219–228. |
| [12] | V. K. Chawla, A. K. Chanda, S. Angra, et al., Multi-load AGVs scheduling by application of modified memetic particle swarm optimization algorithm, J. Braz. Socy. Mech. Sci., 2018. Available from: 10.1007/s40430-018-1357-4. |
| [13] | S. Bakshi, Z. Y. Yan, D. M. Chen, et al., A fast algorithm on minimum-time scheduling of an autonomous ground vehicle using a traveling salesman framework, J. Dyn. Syst. Meas. Control, 2018. Available from: DOI, 10.1115/1.4040665. |
| [14] | H. Fazlollahtabar and S. Hassanli, Hybrid cost and time path planning for multiple autonomous guided vehicles, Appl. Intell., 48 (2018), 482–498. |
| [15] | M. Grunow, H. Gunther, M. Lehmann, et al., Dispatching multi-load AGVs in highly automatedseaport container terminals, OR Spectrum., 26 (2014), 211–235. |
| [16] | T. J. Park, J. W. Ahn, C. S. Han, et al., A path generation algorithm of an automatic guided vehicle using sensor scanning method, J. Mech. Sci. Technol., 16 (2002), 137–146. |
| [17] | H. Y. Wang, Q. Huang, C. T. Li, et al., Graph theory algorithm and MATLAB simulation, BeiJing (2009), Beihang University Press. |
| [18] | A. Ali and M Tawhid, A hybrid particle swarm optimization and genetic algorithm with population partitioning for large scale optimization problems, Ain. Shams. Eng. J., 8 (2017), 191–206. |
| [19] | X. Y. Li and L. Gao, An effective hybrid genetic algorithm and tabu search for flexible job shop scheduling problem, Int. J. Prod. Econ., 174 (2016), 93–110. |
| [20] | P. B. C. Miranda and R. B. C. Prudêncio, Generation of particle swarm optimization algorithms: An experimental study using grammar guided genetic, Appl. Soft. Comput., 60 (2017), 281–296. |
| [21] | S. Hamed and K. Govindan, A hybrid particle swarm optimization and genetic algorithm for closed loop supply chain network design in large scale networks, Appl. Math. Model., 39 (2015), 3990–4012. |
| [22] | K. Deb and N. Padhye, Enhancing performance of particle swarm optimization through an algorithmic link with genetic algorithms, Comput. Optim. Appl., 57 (2014), 761–794. |
| [23] | X. Y. Li, L. Gao, Q. K. Pan, et al., An effective hybrid genetic algorithm and variable neighborhood search for integrated process planning and scheduling in a packaging machine workshop, IEEE T. Syst. Man. Cy. S., 2018. |
| [24] | A. M. Bertram, Q. Zhang, S. C. Kong, et al., A novel particle swarm and genetic algorithm hybrid method for diesel engine performance optimization, Int. J. Engine. Res., 17 (2016), 732–747. |
| [25] | X. Y. Li, C. Lu, L. Gao, et al., An effective multi-objective algorithm for energy efficient scheduling in a real-life welding shop, IEEE T. Ind. Inform., 14 (2018), 5400–5409. |
| [26] | Y. N. Yan, L. Z. Liu, B. Y. Luo, et al., Arrangement of garment production line by particle swarm algorithm, J. Text. Res., 39 (2018), 120–124. |
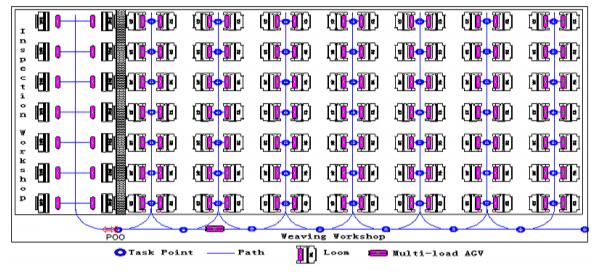
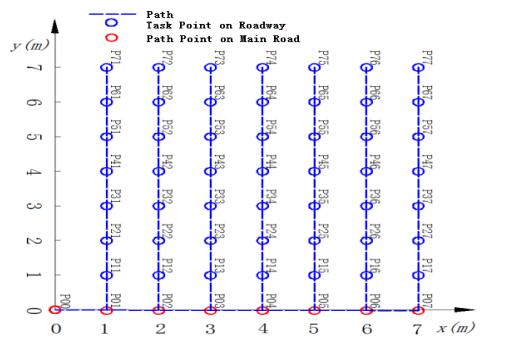
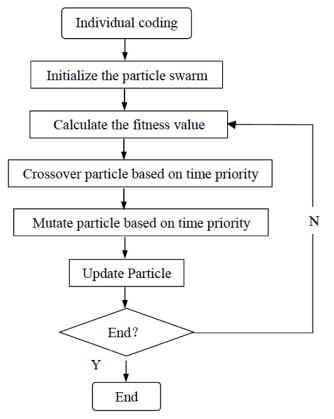
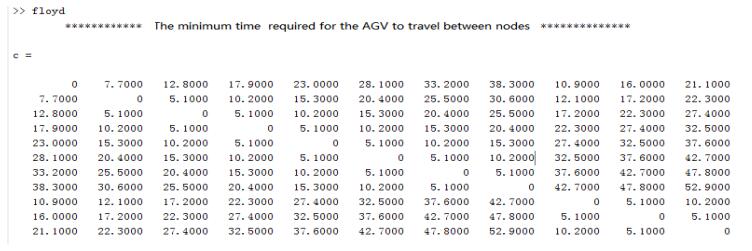

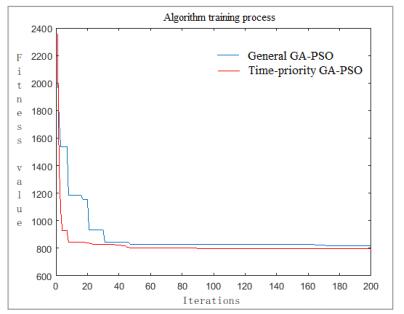
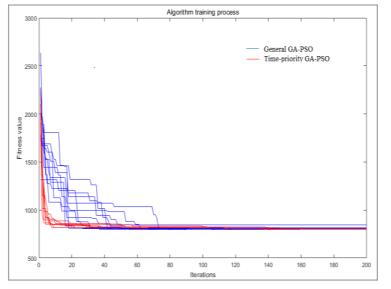
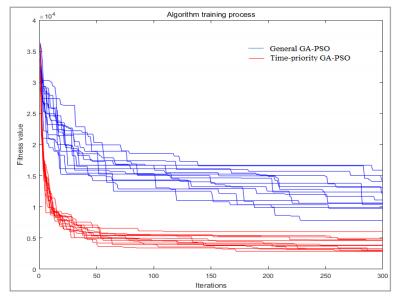
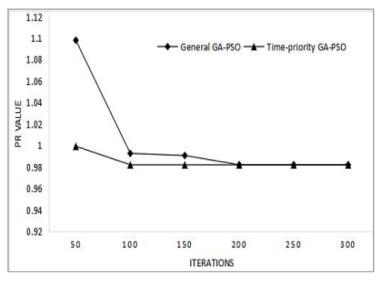
Figures(10) / Tables(4)

Li-zhen Du, Shanfu Ke, Zhen Wang, Jing Tao, Lianqing Yu, Hongjun Li. Research on multi-load AGV path planning of weaving workshop based on time priority[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2277-2292. doi: 10.3934/mbe.2019113







 DownLoad:
DownLoad: