Citation: Qianqian Cui. Global stability of multi-group SIR epidemic model with group mixing and human movement[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 1798-1814. doi: 10.3934/mbe.2019087

| [1] | M. Kermark and A. Mckendrick, Contributions to the mathematical theory of epidemics, Proc. Roy. Soc. A, 115 (1927), 700–721. |
| [2] | H. Shu, D. Fan and J.Wei, Global stability of multi-group SEIR epidemic models with distributed delays and nonlinear transmission, Nonlinear Anal. Real World Appl., 13 (2012), 1581–1592. |
| [3] | H. Guo, M. Li and Z. Shuai, Global stability of the endemic equilibrium of multigroup SIR epidemic models, Canadian Appl. Math. Quart., 14 (2006), 259–284. |
| [4] | H. Guo, M. Y. Li and Z. Shuai, A graph-theoretic approach to the method of global Lyapunov functions, Proc. Amer. Math. Soc., 136 (2008), 2793–2802. |
| [5] | T. Kuniya, Y. Murya and Y. Enatsu, Threshold dynamics of an SIR epidemic model with hybird of nultigroup and patch structures, Math. Biosci. Eng., 11 (2014), 1357–1393. |
| [6] | T. Kuniya and Y. Muroya, Global stability of a multi-group SIS epidemic model with varying total population size, Appl. Math. Comput., 265 (2015), 785–798. |
| [7] | A. Lajmanovich and J. A. York, A deterministic model for gonorrhea in a nonhomogeneous population, Math. Biosci., 28 (1976), 221–236. |
| [8] | M. Y. Li and Z. Shuai, Global stability of an epidemic model in a patchy environment, Canadian Appl. Math. Quart., 17 (2009), 175–187. |
| [9] | M. Y. Li and Z. Shuai, Global-stability problem for coupled systems of differential equations on networks, J. Differ. Eqn., 248 (2010), 1–20. |
| [10] | Y. Muroya, Y. Enatsu and T. Kuniya, Global stability for a multi-group SIRS epidemic model with varying population sizes, Nonlinear Anal. Real World Appl, 14 (2013), 1693–1704. |
| [11] | H. L. Smith, P. Waltman, The Theory of the Chemostat, Cambridge University, 1995. |
| [12] | R. Sun and J. Shi, Global stability of multigroup epidemic model with group mixing and nonlinear incidence rates, Appl. Math. Comput., 218 (2011), 280–286. |
| [13] | W. Wang and X. Zhao, An epidemic model with population dispersal and infection period, SIAM J. Appl. Math., 66 (2006), 1454–1472. |
| [14] | L.Wang andW. Yang, Global dynamics of a two-patch SIS model with infection during transport, Appl. Math. Comput., 217 (2011), 8458-8467. |
| [15] | How Many Ebola Patients Have Been Treated Outside of Africa?, Source of New York Times, 2014. Available from: https://ritholtz.com/2014/10/ how-many-ebola-patients-have-been-treated-outside-africa/. |
| [16] | J. Arino, J. R. Davis, D. Hartley, et al., A multi-species epidemic model with spatial dynamics, Math. Med. Biol., 22 (2005), 129–142. |
| [17] | X. Liu and Y. Takeuchi, Spread of disease with transport-related infection and entry screening, J Theor. Biol., 242 (2006), 517–528. |
| [18] | T. Chen, Z. Sun and B. Wu, Stability of multi-group models with cross-dispersal based on graph theory, Appl. Math. Model., 47 (2017), 745–754. |
| [19] | H. W. Hethcote, The mathematics of infectious diseases, SIAM Rev., 42 (2000), 599–653. |
| [20] | T. Kuniya and Y. Muroya, Global stability of a multi-group SIS epidemic model for population migration, Discrete Continuous Dynam. Systems - B, 19 (2014), 1105–1118. |
| [21] | Y. Muroya, Y. Enatsu and T. Kuniya, Global stability of extended multi-group SIR epidemic models with patches through migration and cross patch infection, Acta Math. Sci., 33 (2013), 341–361. |
| [22] | A. Berman and R. J. Plemmons, Nonnegative Matrices in the Mathematical Sciences, Academic Press, New York, 1979. |
| [23] | N. P. Bhatia and G. P. Szegö, Dynamical Systems: Stability Theory and Applications, Springer, Berlin, 2006. |
| [24] | C. Castillo-Chavez and H. Thieme, Asymptotically autonomous epidemic models, in Mathematical Population Dynamics: Analysis of Heterogeneity (eds. O. Arino, D. Axelrod, M. Kimmel and M. Langlais), Springer, Berlin, 1995, 33–50. |
| [25] | P. van den Driessche and J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transimission, Math. Biosci., 180 (2002), 29–48. |
| [26] | M. C. Eisenberg, Z. Shuai, J. H. Tien, et al., A cholera model in a patchy environment with water and human movement, Math. Biosci., 246 (2013), 105–112. |
| [27] | H. I. Freedman, S. Ruan and M. Tang, Uniform persistence and flows near a closed positively invariant set, J. Dyn. Differ. Equ., 6 (1994), 583–600. |
| [28] | K. Mischaikow, H. L. Smith and H. R. Thieme, Asymptotically autonomous semiflows: Chain recur-rence and Lyapunov functions, Trans. Am. Math. Soc., 347 (1995), 1669–1685. |
| [29] | J. P. LaSalle and S. Kefscgetz, Stability by Liapunovs Direct Method with Applications, Academic Press, New York, 1961. |
| [30] | M. Y. Li, J. R. Graef, L. Wang, et al., Global dynamics of a SEIR model with varying total population size, Math. Biosci., 160 (1999), 191–213. |
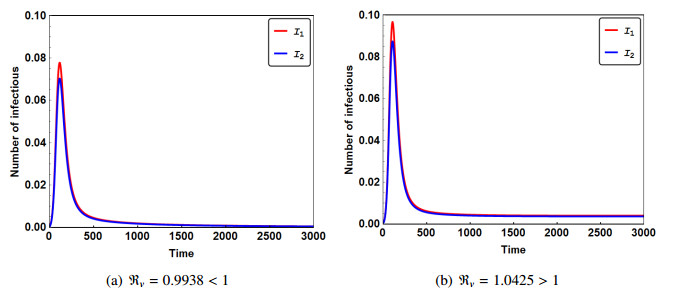
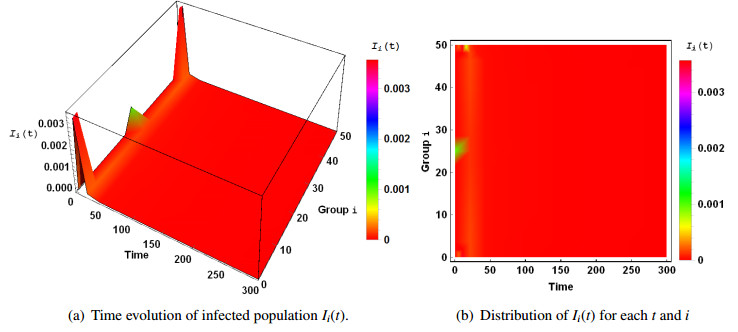
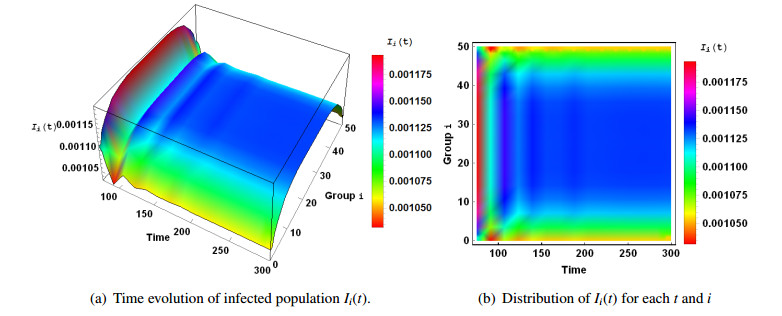
Figures(3)

Qianqian Cui. Global stability of multi-group SIR epidemic model with group mixing and human movement[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 1798-1814. doi: 10.3934/mbe.2019087







 DownLoad:
DownLoad: