Citation: Xiaoying Wang, Xingfu Zou. Pattern formation of a predator-prey model with the cost of anti-predator behaviors[J]. Mathematical Biosciences and Engineering, 2018, 15(3): 775-805. doi: 10.3934/mbe.2018035

| [1] | [ B. E. Ainseba,M. Bendahmane,A. Noussair, A reaction-diffusion system modeling predator-prey with prey-taxis, Nonlinear Analysis: Real World Applications, 9 (2008): 2086-2105. |
| [2] | [ N. D. Alikakos, An application of the invariance principle to reaction-diffusion equations, Journal of Differential Equations, 33 (1979): 201-225. |
| [3] | [ D. Alonso,F. Bartumeus,J. Catalan, Mutual interference between predators can give rise to Turing spatial patterns, Ecology, 83 (2002): 28-34. |
| [4] | [ H. Amann, Dynamic theory of quasilinear parabolic equations Ⅱ: Reaction-diffusion systems, Differential Integral Equations, 3 (1990): 13-75. |
| [5] | [ H. Amann, Dynamic theory of quasilinear parabolic systems Ⅲ: Global existence, Mathematische Zeitschrift, 202 (1989): 219-250. |
| [6] | [ H. Amann, Nonhomogeneous linear and quasilinear elliptic and parabolic boundary value problems, Function Spaces, Differential Operators and Nonlinear Analysis, 133 (1993): 9-126. |
| [7] | [ J. R. Beddington, Mutual interference between parasites or predators and its effect on searching efficiency, Journal of Animal Ecology, 44 (1975): 331-340. |
| [8] | [ V. N. Biktashev,J. Brindley,A. V. Holden,M. A. Tsyganov, Pursuit-evasion predator-prey waves in two spatial dimensions, Chaos: An Interdisciplinary Journal of Nonlinear Science, 14 (2004): 988-994. |
| [9] | [ X. Cao, Y. Song and T. Zhang, Hopf bifurcation and delay-induced Turing instability in a diffusive lac operon model, International Journal of Bifurcation and Chaos, 26 (2016), 1650167, 22pp. |
| [10] | [ S. Creel,D. Christianson, Relationships between direct predation and risk effects, Trends in Ecology & Evolution, 23 (2008): 194-201. |
| [11] | [ W. Cresswell, Predation in bird populations, Journal of Ornithology, 152 (2011): 251-263. |
| [12] | [ D. L. DeAngelis,R. A. Goldstein,R. V. O'neill, A model for tropic interaction, Ecology, 56 (1975): 881-892. |
| [13] | [ T. Hillen,K. J. Painter, A user's guide to PDE models for chemotaxis, Journal of Mathematical Biology, 58 (2009): 183-217. |
| [14] | [ T. Hillen,K. J. Painter, Global existence for a parabolic chemotaxis model with prevention of overcrowding, Advances in Applied Mathematics, 26 (2001): 280-301. |
| [15] | [ C. S. Holling, The components of predation as revealed by a study of small-mammal predation of the European pine sawfly, The Canadian Entomologist, 91 (1959): 293-320. |
| [16] | [ C. S. Holling, Some characteristics of simple types of predation and parasitism, The Canadian Entomologist, 91 (1959): 385-398. |
| [17] | [ H. Jin,Z. Wang, Global stability of prey-taxis systems, Journal of Differential Equations, 262 (2017): 1257-1290. |
| [18] | [ J. M. Lee,T. Hillen,M. A. Lewis, Pattern formation in prey-taxis systems, Journal of Biological Dynamics, 3 (2009): 551-573. |
| [19] | [ S. A. Levin, The problem of pattern and scale in ecology, Ecology, 73 (1992): 1943-1967. |
| [20] | [ E. A. McGehee,E. Peacock-López, Turing patterns in a modified Lotka-Volterra model, Physics Letters A, 342 (2005): 90-98. |
| [21] | [ A. B. Medvinsky,S. V. Petrovskii,I. A. Tikhonova,H. Malchow,B. Li, Spatiotemporal complexity of plankton and fish dynamics, SIAM Review, 44 (2002): 311-370. |
| [22] | [ J. D. Meiss, Differential Dynamical Systems, SIAM, 2007. |
| [23] | [ M. Mimura,J. D. Murray, On a diffusive prey-predator model which exhibits patchiness, Journal of Theoretical Biology, 75 (1978): 249-262. |
| [24] | [ M. Mimura, Asymptotic behavior of a parabolic system related to a planktonic prey and predator system, SIAM Journal on Applied Mathematics, 37 (1979): 499-512. |
| [25] | [ A. Morozov,S. Petrovskii,B. Li, Spatiotemporal complexity of patchy invasion in a predator-prey system with the Allee effect, Journal of Theoretical Biology, 238 (2006): 18-35. |
| [26] | [ J. D. Murray, Mathematical Biology Ⅱ: Spatial Models and Biomedical Applications, Third edition. Interdisciplinary Applied Mathematics, 18. Springer-Verlag, New York, 2003. |
| [27] | [ W. Ni,M. Wang, Dynamics and patterns of a diffusive Leslie-Gower prey-predator model with strong Allee effect in prey, Journal of Differential Equations, 261 (2016): 4244-4274. |
| [28] | [ K. J. Painter,T. Hillen, Volume-filling and quorum-sensing in models for chemosensitive movement, Can. Appl. Math. Quart, 10 (2002): 501-543. |
| [29] | [ S. V. Petrovskii,A. Y. Morozov,E. Venturino, Allee effect makes possible patchy invasion in a predator-prey system, Ecology Letters, 5 (2002): 345-352. |
| [30] | [ D. Ryan,R. Cantrell, Avoidance behavior in intraguild predation communities: A cross-diffusion model, Discrete and Continuous Dynamical Systems, 35 (2015): 1641-1663. |
| [31] | [ H. Shi,S. Ruan, Spatial, temporal and spatiotemporal patterns of diffusive predator-prey models with mutual interference, IMA Journal of Applied Mathematics, 80 (2015): 1534-1568. |
| [32] | [ H. L. Smith, Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems, Mathematical Surveys and Monographs, 41. American Mathematical Society, Providence, RI, 1995. |
| [33] | [ Y. Song,X. Zou, Spatiotemporal dynamics in a diffusive ratio-dependent predator-prey model near a Hopf-Turing bifurcation point, Computers and Mathematics with Applications, 37 (2014): 1978-1997. |
| [34] | [ Y. Song,T. Zhang,Y. Peng, Turing-Hopf bifurcation in the reaction-diffusion equations and its applications, Communications in Nonlinear Science and Numerical Simulation, 33 (2016): 229-258. |
| [35] | [ Y. Song and X. Tang, Stability, steady-state bifurcations, and Turing patterns in a predator-prey model with herd behavior and prey-taxis, Studies in Applied Mathematics,(2017). |
| [36] | [ J. H. Steele, Spatial Pattern in Plankton Communities (Vol. 3), Springer Science & Business Media, 1978. |
| [37] | [ Y. Tao, Global existence of classical solutions to a predator-prey model with nolinear prey-taxis, Nonlinear Analysis: Real World Applications, 11 (2010): 2056-2064. |
| [38] | [ J. Wang,J. Shi,J. Wei, Dynamics and pattern formation in a diffusive predator-prey system with strong Allee effect in prey, Journal of Differential Equations, 251 (2011): 1276-1304. |
| [39] | [ J. Wang,J. Wei,J. Shi, Global bifurcation analysis and pattern formation in homogeneous diffusive predator-prey systems, Journal of Differential Equations, 260 (2016): 3495-3523. |
| [40] | [ X. Wang,L. Y. Zanette,X. Zou, Modelling the fear effect in predator-prey interactions, Journal of Mathematical Biology, 73 (2016): 1179-1204. |
| [41] | [ X. Wang,W. Wang,G. Zhang, Global bifurcation of solutions for a predator-prey model with prey-taxis, Mathematical Methods in the Applied Sciences, 38 (2015): 431-443. |
| [42] | [ Z. Wang and T. Hillen, Classical solutions and pattern formation for a volume filling chemotaxis model, Chaos: An Interdisciplinary Journal of Nonlinear Science, 17 (2007), 037108, 13 pp. |
| [43] | [ S. Wu,J. Shi,B. Wu, Global existence of solutions and uniform persistence of a diffusive predator-prey model with prey-taxis, Journal of Differential Equations, 260 (2016): 5847-5874. |
| [44] | [ J. Xu,G. Yang,H. Xi,J. Su, Pattern dynamics of a predator-prey reaction-diffusion model with spatiotemporal delay, Nonlinear Dynamics, 81 (2015): 2155-2163. |
| [45] | [ F. Yi,J. Wei,J. Shi, Bifurcation and spatiotemporal patterns in a homogeneous diffusive predator-prey system, Journal of Differential Equations, 246 (2009): 1944-1977. |
| [46] | [ L. Y. Zanette,A. F. White,M. C. Allen,M. Clinchy, Perceived predation risk reduces the number of offspring songbirds produce per year, Science, 334 (2011): 1398-1401. |
| [47] | [ T. Zhang and H. Zang, Delay-induced Turing instability in reaction-diffusion equations, Physical Review E, 90 (2014), 052908. |
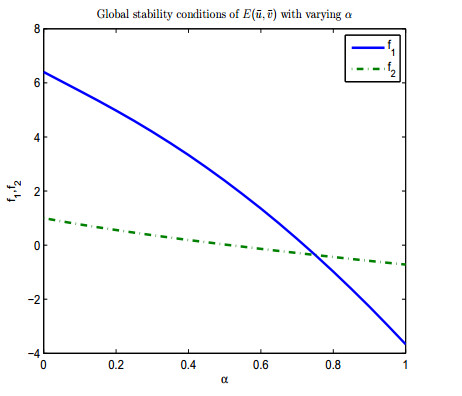
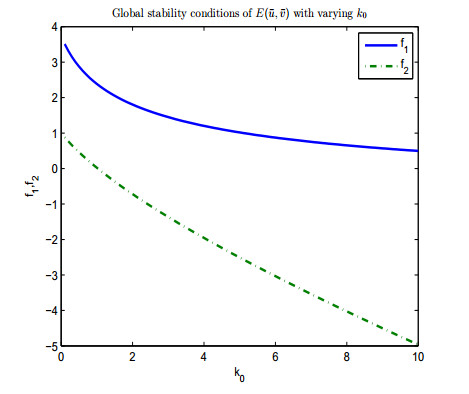
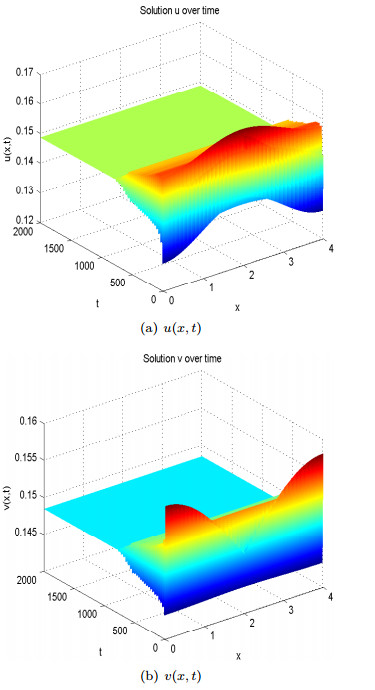
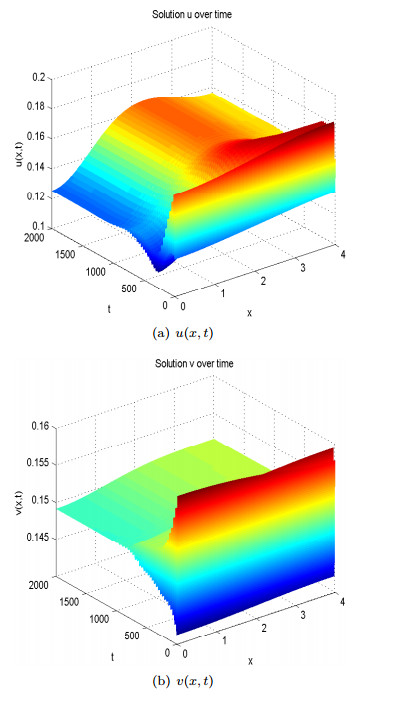
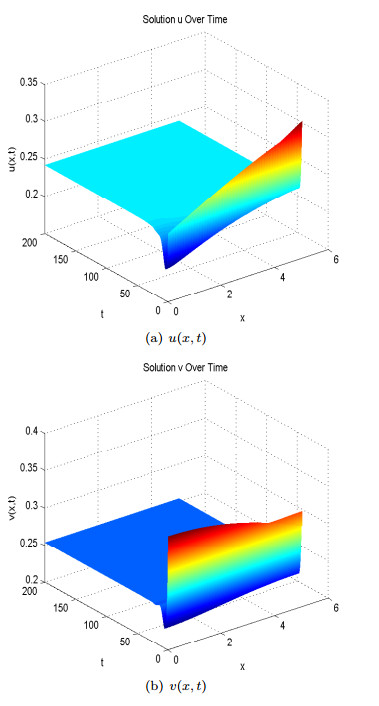
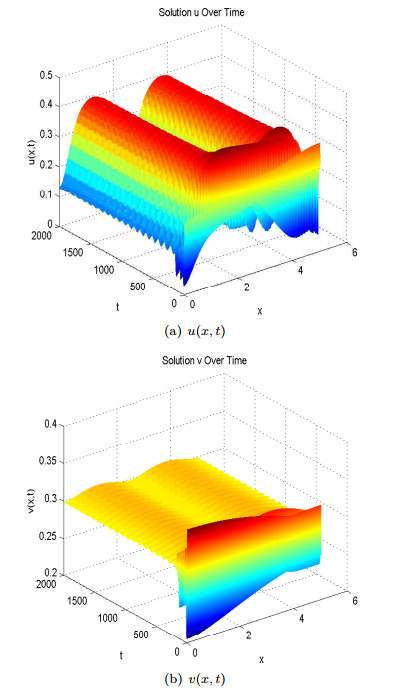
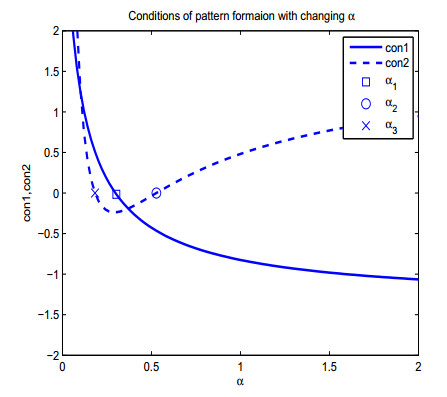
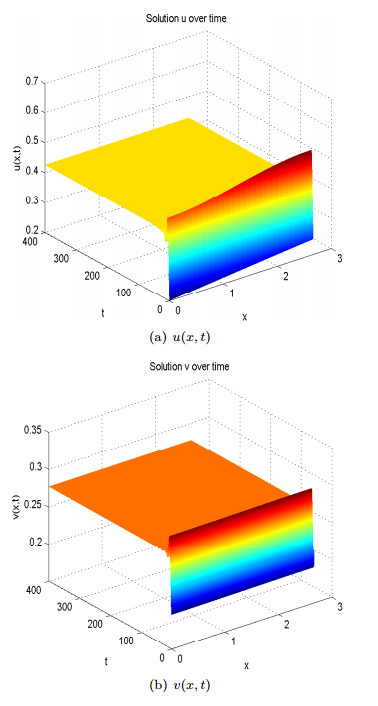
Figures(13)

Xiaoying Wang, Xingfu Zou. Pattern formation of a predator-prey model with the cost of anti-predator behaviors[J]. Mathematical Biosciences and Engineering, 2018, 15(3): 775-805. doi: 10.3934/mbe.2018035







 DownLoad:
DownLoad: