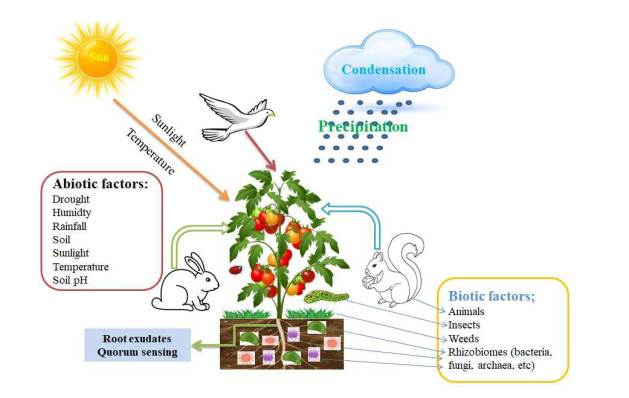
Plant microbiota has a variety of impacts on the plant. Some are beneficial, while some are pathogenic. This study discusses the general metagenomics procedures in processing plant-related metagenomes and focuses on the tomato plants' rhizosphere species. Metagenomics, associated with eventual DNA, is isolated from environmental samples and thus permits absolute microbial population identification. Meanwhile, the genetic content of the DNA sample obtained allows the functional capability identification and biochemical procedure of many microorganisms. This review reveals the recent utilization and application of the potential of Next-Generation Sequencing (NGS) in agriculture. It involves plant-associated microbiota, the factors driving their diversity, and plant metagenome to tackle current challenges experienced in food security. This review provides the newest methods for rapidly identifying the microbial communities inhabiting the rhizosphere soil of tomato plants.
Citation: Afeez Adesina Adedayo, Samir Ben Romdhane, Olubukola Oluranti Babalola. Metagenomic study of the tomato rhizosphere soil microbiome: current state and prospect[J]. AIMS Geosciences, 2023, 9(2): 330-356. doi: 10.3934/geosci.2023018
Plant microbiota has a variety of impacts on the plant. Some are beneficial, while some are pathogenic. This study discusses the general metagenomics procedures in processing plant-related metagenomes and focuses on the tomato plants' rhizosphere species. Metagenomics, associated with eventual DNA, is isolated from environmental samples and thus permits absolute microbial population identification. Meanwhile, the genetic content of the DNA sample obtained allows the functional capability identification and biochemical procedure of many microorganisms. This review reveals the recent utilization and application of the potential of Next-Generation Sequencing (NGS) in agriculture. It involves plant-associated microbiota, the factors driving their diversity, and plant metagenome to tackle current challenges experienced in food security. This review provides the newest methods for rapidly identifying the microbial communities inhabiting the rhizosphere soil of tomato plants.

| [1] |
Ahmed N, Turchini GM (2021) The evolution of the Blue-Green revolution of Rice-Fish cultivation for Sustainable Food Production. Sustain Sci 16: 1375–1390. https://doi.org/10.1007/s11625-021-00924-z doi: 10.1007/s11625-021-00924-z

|
| [2] |
Phillips FC, Ceesay EK (2021) Livelihood Dependence and Forest Reserve Management in Ijaiye Forest Reserve, Oyo State, Nigeria. Int J For Hortic 6: 26–39. http://dx.doi.org/10.20431/2454-9487.0602003 doi: 10.20431/2454-9487.0602003
|
| [3] |
Adedayo AA, Fadiji AE, Babalola OO (2022) The Effects of Plant Health Status on the Community Structure and Metabolic Pathways of Rhizosphere Microbial Communities Associated with Solanum lycopersicum. Horticulturae 8: 404. https://dx.doi.org/10.3390/horticulturae8050404 doi: 10.3390/horticulturae8050404
|
| [4] |
Vanlerberghe GC (2013) Alternative Oxidase: A Mitochondrial Respiratory Pathway to Maintain Metabolic and Signaling Homeostasis during Abiotic and Biotic Stress in Plants. Int J Mol Sci 14: 6805–6847. http://dx.doi.org/10.3390/ijms14046805 doi: 10.3390/ijms14046805
|
| [5] |
Sachdev S, Bauddh K, Singh RP (2023) Native Rhizospheric Microbes Mediated Management of Biotic Stress and Growth Promotion of Tomato. Sustainability 15: 593. https://doi.org/10.3390/su15010593 doi: 10.3390/su15010593
|
| [6] |
Yngstrom I (2002) Women, Wives and Land Rights in Africa: Situating Gender Beyond the Household in the Debate Over Land Policy and Changing Tenure Systems. Oxford Dev Stud 30: 21–40. http://dx.doi.org/10.1080/136008101200114886 doi: 10.1080/136008101200114886
|
| [7] |
Kumar A, Vyas P, Malla MA, et al. (2019) Taxonomic and Functional Annotation of Termite Degraded Butea monosperma (Lam.) Kuntze (Flame of the Forest). Open Microbiol J 13: 154–163. http://dx.doi.org/10.2174/1874285801913010154 doi: 10.2174/1874285801913010154
|
| [8] |
Yadav U, Bano N, Bag S, et al. (2022) An Insight into the Endophytic Bacterial Community of Tomato after Spray Application of Propiconazole and Bacillus subtilis Strain NBRI-W9. Microbiol Spectrum 10: e01186–01122. http://dx.doi.org/10.1128/spectrum.01186-22 doi: 10.1128/spectrum.01186-22
|
| [9] |
Compant S, Samad A, Faist H, et al. (2019) A Review on the Plant Microbiome: Ecology, Functions, and emerging trends in Microbial Application. J Adv Res 19: 29–37. http://dx.doi.org/10.1016/j.jare.2019.03.004 doi: 10.1016/j.jare.2019.03.004
|
| [10] |
Liu H, Brettell LE, Singh B (2020) Linking the Phyllosphere Microbiome to Plant Health. Trends Plant Sci 25: 841–844. http://dx.doi.org/10.1016/j.tplants.2020.06.003 doi: 10.1016/j.tplants.2020.06.003
|
| [11] |
Shimp JF, Tracy JC, Davis LC, et al. (1993) Beneficial effects of plants in the remediation of soil and groundwater contaminated with organic materials. Crit Rev Environ Sci Technol. 23: 41–77. http://dx.doi.org/10.1080/10643389309388441 doi: 10.1080/10643389309388441
|
| [12] | Nihorimbere V, Ongena M, Smargiassi M, et al. (2011) Beneficial effect of the rhizosphere microbial community for plant growth and health. Biotechnol Agron Soc Environ 15: 327–337. |
| [13] |
Alexander HM (2010) Disease in natural plant populations, communities, and ecosystems: insights into ecological and evolutionary processes. Plant Dis 94: 492–503. http://dx.doi.org/10.1094/PDIS-94-5-0492 doi: 10.1094/PDIS-94-5-0492
|
| [14] |
Sugimoto K, Zager JJ, Aubin BS, et al. (2022) Flavonoid deficiency disrupts redox homeostasis and terpenoid biosynthesis in glandular trichomes of tomato. Plant Physiol 188: 1450–1468. https://doi.org/10.1093/plphys/kiab488 doi: 10.1093/plphys/kiab488
|
| [15] |
Jones P, Benjamin J. Garcia, Furches A, et al. (2019) Plant Host-Associated Mechanisms for Microbial Selection. Front Plant Sci 10: 1–14. https://doi.org/10.3389/fpls.2019.00862 doi: 10.3389/fpls.2019.00862
|
| [16] |
Odelade K, Babalola O (2019) Bacteria, Fungi and Archaea Domains in Rhizospheric Soil and their effects in enhancing Agricultural Productivity. Int J Environ Res Public Health 16: 3873. https://doi.org/10.3390/ijerph16203873 doi: 10.3390/ijerph16203873
|
| [17] |
Kumar A, Dubey A (2020) Rhizosphere microbiome: Engineering bacterial competitiveness for enhancing crop production. J Adv Res 24: 337–352. https://doi.org/10.1016/j.jare.2020.04.014 doi: 10.1016/j.jare.2020.04.014
|
| [18] |
Levy A, Conway JM, Dangl JL, et al. (2018) Elucidating Bacterial Gene Functions in the Plant Microbiome. Cell Host Microbe 24: 475–485. https://doi.org/10.1016/j.chom.2018.09.005 doi: 10.1016/j.chom.2018.09.005
|
| [19] |
Park JM, Hong JW, Lee W, et al. (2021) Geographical Isolation and Root-Associated Fungi in the Marine Terrains: A Step Toward Establishing a Strategy for Acquiring Unique Microbial Resources. Mycobiology 49: 235–248. https://doi.org/10.1080/12298093.2021.1913826 doi: 10.1080/12298093.2021.1913826
|
| [20] |
Hassani MA, Durán P, Hacquard S (2018) Microbial interactions within the plant holobiont. Microbiome 6: 1–17. https://doi.org/10.1186/s40168-018-0445-0 doi: 10.1186/s40168-018-0445-0
|
| [21] |
Adedayo AA, Babalola OO, Prigent-Combaret C, et al. (2022) The application of plant growth-promoting rhizobacteria in Solanum lycopersicum production in the agricultural system: a review. PeerJ 10: e13405. https://doi.org/10.7717/peerj.13405 doi: 10.7717/peerj.13405
|
| [22] |
Fadiji AE, Babalola OO (2020) Elucidating Mechanisms of Endophytes used in Plant Protection and other Bioactivities With Multifunctional Prospects. Front Bioeng Biotechnol 8: 1–20. https://doi.org/10.3389/fbioe.2020.00467 doi: 10.3389/fbioe.2020.00467
|
| [23] |
Bai Y, Kissoudis C, Yan Z, et al. (2018) Plant behaviour under combined stress: Tomato responses to combined salinity and pathogen stress. Plant J 93: 781–793. https://doi.org/10.1111/tpj.13800 doi: 10.1111/tpj.13800
|
| [24] |
Leneveu-Jenvrin C, Charles F, Barba FJ, et al. (2020) Role of biological control agents and physical treatments in maintaining the quality of fresh and minimally-processed fruit and vegetables. Crit Rev Food Sci Nutr 60: 2837–2855. https://doi.org/10.1080/10408398.2019.1664979 doi: 10.1080/10408398.2019.1664979
|
| [25] |
Constantin ME, de Lamo FJ, Vlieger BV, et al. (2019) Endophyte-Mediated Resistance in Tomato to Fusarium oxysporum is Independent of ET, JA, and SA. Front Plant Sci 10: 1–14. https://doi.org/10.3389/fpls.2019.00979 doi: 10.3389/fpls.2019.00979
|
| [26] |
Cheng Z, Lei S, Li Y, et al. (2020) Revealing the Variation and Stability of Bacterial Communities in Tomato Rhizosphere Microbiota. Microorganisms 8: 170. https://doi.org/10.3390/microorganisms8020170 doi: 10.3390/microorganisms8020170
|
| [27] |
Azeem M, Soundari PG, Ali A, et al. (2021) Soil metaphenomics: a step forward in metagenomics. Arch Agron Soil Sci: 1–19. https://doi.org/10.1080/03650340.2021.1921743 doi: 10.1080/03650340.2021.1921743
|
| [28] |
Odelade KA, Babalola OO (2019) Bacteria, Fungi and Archaea Domains in Rhizospheric Soil and their effects in enhancing Agricultural Productivity. Int J Environ Res Public Health 16: 3873. https://doi.org/10.3390/ijerph16203873 doi: 10.3390/ijerph16203873
|
| [29] |
Nwachukwu BC, Ayangbenro AS, Babalola OO (2021) Elucidating the Rhizosphere Associated Bacteria for Environmental Sustainability. Agriculture 11: 75. https://doi.org/10.3390/agriculture11010075 doi: 10.3390/agriculture11010075
|
| [30] |
Backer R, Rokem JS, Ilangumaran G, et al. (2018) Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front Plant Sci 9: 1–17. https://doi.org/10.3389/fpls.2018.01473 doi: 10.3389/fpls.2018.01473
|
| [31] |
Semchenko M, Barry KE, Vries FT, et al. (2022) Deciphering the role of specialist and generalist plant–microbial interactions as drivers of plant–soil feedback. New Phytol 234: 1929–1944. https://doi.org/10.1111/nph.18118 doi: 10.1111/nph.18118
|
| [32] |
Saeed Q, Xiukang W, Haider FU, et al. (2021) Rhizosphere Bacteria in Plant Growth Promotion, Biocontrol, and Bioremediation of Contaminated Sites: A Comprehensive Review of Effects and Mechanisms. Int J Mol Sci 22: 10529. https://doi.org/10.3390/ijms221910529 doi: 10.3390/ijms221910529
|
| [33] |
Sokol NW, Slessarev E, Marschmann GL, et al. (2022) Life and death in the soil microbiome: how ecological processes influence biogeochemistry. Nat Rev Microbiol 20: 415–430. https://doi.org/10.1038/s41579-022-00695-z doi: 10.1038/s41579-022-00695-z
|
| [34] |
Long XE, Yao H, Huang Y, et al. (2018) Phosphate levels influence the utilisation of Rice Rhizodeposition Carbon and the Phosphate-Solubilising Microbial Community in a paddy soil. Soil Biol Biochem 118: 103–114. https://doi.org/10.1016/j.soilbio.2017.12.014 doi: 10.1016/j.soilbio.2017.12.014
|
| [35] |
Qu Q, Zhang Z, Peijnenburg WJGM, et al. (2020) Rhizosphere Microbiome Assembly and Its Impact on Plant Growth. J Agric Food Chem 68: 5024–5038. https://doi.org/10.1021/acs.jafc.0c00073 doi: 10.1021/acs.jafc.0c00073
|
| [36] | Rani SG, Kumari H, Pati P, et al. (2021) Recent approaches of systems Biology and Omics in Plant Research. J Curr Opin Crop Sci 2: 288–305. |
| [37] |
Adeleke BS, Babalola OO (2021) The endosphere microbial communities, a great promise in agriculture. Int Microbiol 24: 1–17. https://doi.org/10.1007/s10123-020-00140-2 doi: 10.1007/s10123-020-00140-2
|
| [38] | Mukherjee S, Bassler BL (2019) Bacterial Quorum Sensing in complex and dynamically changing environments. Nat Rev Microbiol 17: 371–382 |
| [39] |
Krzyżek P (2019) Challenges and Limitations of Anti-quorum Sensing Therapies. Front Microbiol 10: 1–10. https://doi.org/10.3389/fmicb.2019.02473 doi: 10.3389/fmicb.2019.02473
|
| [40] |
Pena RT, Blasco L, Ambroa A, et al. (2019) Relationship Between Quorum Sensing and Secretion Systems. Front Microbiol 10: 1–14. https://doi.org/10.3389/fmicb.2019.01100 doi: 10.3389/fmicb.2019.01100
|
| [41] |
Walters WA, Jin Z, Youngblut N, et al. (2018) Large-scale replicated Field study of Maize Rhizosphere identifies heritable microbes. Proc Natl Acad Sci U S A 115: 7368–7373. https://doi.org/10.1073/pnas.1800918115 doi: 10.1073/pnas.1800918115
|
| [42] |
De Corato U (2020) Soil Microbiota manipulation and its role in suppressing Soil-Borne Plant Pathogens in Organic Farming Systems under the light of Microbiome-Assisted Strategies. Chem Biol Technol Agric 7: 1–26. https://doi.org/10.1186/s40538-020-00183-7 doi: 10.1186/s40538-020-00183-7
|
| [43] |
Timmis K, Ramos JL (2021) The soil crisis: The need to treat as a Global Health Problem and the pivotal role of Microbes in Prophylaxis and Therapy. Microb Biotechnol 14: 769–797. https://doi.org/10.1111/1751-7915.13771 doi: 10.1111/1751-7915.13771
|
| [44] |
Tosif MM, Najda A, Bains A, et al. (2021) A Comprehensive Review on Plant-Derived Mucilage: Characterization, Functional Properties, Applications, and Its Utilization for Nanocarrier Fabrication. Polymers 13: 1066. https://doi.org/10.3390/polym13071066 doi: 10.3390/polym13071066
|
| [45] |
Vannette RL (2020) The Floral Microbiome: Plant, Pollinator, and Microbial Perspectives. Annu Rev Ecol Evol Syst 51: 363–386. https://doi.org/10.1146/annurev-ecolsys-011720-013401 doi: 10.1146/annurev-ecolsys-011720-013401
|
| [46] |
Kolbe AR, Aira M, Gómez-Brandón M, et al. (2019) Bacterial succession and functional diversity during vermicomposting of the white grape marc Vitis vinifera v. Albariño. Sci Rep 9: 7472. https://doi.org/10.1038/s41598-019-43907-y doi: 10.1038/s41598-019-43907-y
|
| [47] |
Hajlaoui H, Arraouadi S, Noumi E, et al. (2021) Antimicrobial, Antioxidant, Anti-Acetylcholinesterase, Antidiabetic, and Pharmacokinetic Properties of Carum carvi L. and Coriandrum sativum L. Essential Oils Alone and in Combination. Molecules 26: 3625. https://doi.org/10.3390/molecules26123625 doi: 10.3390/molecules26123625
|
| [48] |
Finlay RD, Mahmood S, Rosenstock N, et al. (2020) Reviews and syntheses: Biological weathering and its consequences at different spatial levels—from nanoscale to global scale. Biogeosciences 17: 1507–1533. https://doi.org/10.5194/bg-17-1507-2020 doi: 10.5194/bg-17-1507-2020
|
| [49] |
Ou Y, Penton CR, Geisen S, et al. (2019) Deciphering Underlying Drivers of Disease Suppressiveness against Pathogenic Fusarium oxysporum. Front Microbiol 10. https://doi.org/10.3389/fmicb.2019.02535 doi: 10.3389/fmicb.2019.02535
|
| [50] |
Wang J, Liu X, Li Y, et al. (2019) Microplastics as contaminants in the soil environment: A mini-review. Sci Total Environ 691: 848–857. https://doi.org/10.1016/j.scitotenv.2019.07.209 doi: 10.1016/j.scitotenv.2019.07.209
|
| [51] |
Wang T, Teng K, Cao Y, et al. (2020) Effects of Lactobacillus hilgardii 60TS-2, with or without homofermentative Lactobacillus plantarum B90, on the Aerobic Stability, Fermentation Quality and Microbial Community dynamics in Sugarcane Top Silage. Bioresour Technol 312: 123600. https://doi.org/10.1016/j.biortech.2020.123600 doi: 10.1016/j.biortech.2020.123600
|
| [52] |
Zhou Y, Watts SE, Boutton TW, et al. (2019) Root Density Distribution and Biomass Allocation of co-occurring Woody Plants on contrasting Soils in a Subtropical Savanna Parkland. Plant Soil 438: 263–279. https://doi.org/10.1007/s11104-019-04018-9 doi: 10.1007/s11104-019-04018-9
|
| [53] |
Mwangangi IM, Büchi L, Haefele SM, et al. (2021) Combining host plant defence with targeted nutrition: key to durable control of hemiparasitic Striga in cereals in sub‐Saharan Africa? New Phytol 230: 2164–2178. https://doi.org/10.1111/nph.17271 doi: 10.1111/nph.17271
|
| [54] |
Lesmes-Vesga RA, Cano LM, Ritenour MA, et al. (2022) Rootstocks for Commercial Peach Production in the Southeastern United States: Current Research, Challenges, and Opportunities. Horticulturae 8: 602. https://doi.org/10.3390/horticulturae8070602 doi: 10.3390/horticulturae8070602
|
| [55] |
De Corato U (2020) Disease-suppressive compost enhances natural soil suppressiveness against soil-borne plant pathogens: A critical review. Rhizosphere 13: 100192. https://doi.org/10.1016/j.rhisph.2020.100192 doi: 10.1016/j.rhisph.2020.100192
|
| [56] |
Toju H, Peay KG, Yamamichi M, et al. (2018) Core Microbiomes for Sustainable Agroecosystems. Nat Plants 4: 247–257. https://doi.org/10.1038/s41477-018-0139-4 doi: 10.1038/s41477-018-0139-4
|
| [57] |
De Corato U (2020) Soil microbiota manipulation and its role in suppressing soil-borne plant pathogens in organic farming systems under the light of microbiome-assisted strategies. Chem Biol Technol Agric 7: 1–26. https://doi.org/10.1186/s40538-020-00183-7 doi: 10.1186/s40538-020-00183-7
|
| [58] |
Sugihara S, Kawashita T, Shitindi M, et al. (2021) Dynamics of fractionated Rhizosphere Soil P and plant P uptake under Maize/P-mobilizing legumes intercropping in strongly weathered soil of Tanzania. Soil Sci Plant Nutr: 1–11. https://doi.org/10.1080/00380768.2021.1911589 doi: 10.1080/00380768.2021.1911589
|
| [59] |
Kalayu G (2019) Phosphate Solubilizing Microorganisms: Promising Approach as Biofertilizers. Int J Agron 2019: 1–7. https://doi.org/10.1155/2019/4917256 doi: 10.1155/2019/4917256
|
| [60] |
Nishigaki T, Sugihara S, Kobayashi K, et al. (2018) Fractionation of Phosphorus in Soils with different Geological and Soil Physicochemical properties in Southern Tanzania. Soil Sci Plant Nutr 64: 291–299. https://doi.org/10.1080/00380768.2018.1436406 doi: 10.1080/00380768.2018.1436406
|
| [61] |
Trivedi P, Mattupalli C, Eversole K, et al. (2021) Enabling sustainable agriculture through understanding and enhancement of microbiomes. New Phytol 230: 2129–2147. https://doi.org/10.1111/nph.17319 doi: 10.1111/nph.17319
|
| [62] |
Akinola SA, Babalola OO (2021) The fungal and archaeal community within plant rhizosphere: a review on their contribution to crop safety. J Plant Nutr 44: 600–618. https://dx.doi.org/10.1080/01904167.2020.1845376 doi: 10.1080/01904167.2020.1845376
|
| [63] |
Taffner J, Erlacher A, Bragina A, et al. (2018) What is the role of Archaea in plants? New insights from the vegetation of alpine bogs. MSphere 3: e00122–00118. https://doi.org/10.1128/mSphere.00122-18 doi: 10.1128/mSphere.00122-18
|
| [64] |
Igiehon NO, Babalola OO (2018) Below-ground-above-ground Plant-microbial Interactions: Focusing on Soybean, Rhizobacteria and Mycorrhizal Fungi. Open Microbiol J 12: 261–279. https://dx.doi.org/10.2174/1874285801812010261 doi: 10.2174/1874285801812010261
|
| [65] |
Dohrmann AB, Küting M, Jünemann S, et al. (2013) Importance of rare taxa for bacterial diversity in the rhizosphere of Bt- and conventional maize varieties. The ISME Journal 7: 37–49. https://doi.org/10.1038/ismej.2012.77 doi: 10.1038/ismej.2012.77
|
| [66] |
Adedayo AA, Fadiji AE, Babalola OO (2022) Plant Health Status Affects the Functional Diversity of the Rhizosphere Microbiome Associated With Solanum lycopersicum. Front Sustain Food Systs 6: 1–10. https://doi.org/10.3389/fsufs.2022.894312 doi: 10.3389/fsufs.2022.894312
|
| [67] |
Shen RF, Cai H, Gong WH (2006) Transgenic Bt cotton has no apparent effect on enzymatic activities or functional diversity of microbial communities in rhizosphere soil. Plant Soil 285: 149–159. https://doi.org/10.1007/s11104-006-9000-z doi: 10.1007/s11104-006-9000-z
|
| [68] |
Kamutando CN, Vikram S, Kamgan-Nkuekam G, et al. (2017) Soil nutritional status and biogeography influence rhizosphere microbial communities associated with the invasive tree Acacia dealbata. Sci Rep 7: 6472. https://doi.org/10.1038/s41598-017-07018-w doi: 10.1038/s41598-017-07018-w
|
| [69] |
Meena MMM, Dubey M, Swapnil PSP, et al. (2017) The rhizosphere microbial community and methods of its analysis. Advances in PGPR research, 275–295. https://doi.org/10.1079/9781786390325.0275 doi: 10.1079/9781786390325.0275
|
| [70] |
Gashaw T, Sitotaw B, Yilma S (2022) Evaluation of Rhizosphere Bacterial Antagonists against Ralstonia solanacearum causing Tomato (Lycopersicon esculentum) Wilt in Central Ethiopia. Int J Agron 2022: 6341555. https://doi.org/10.1155/2022/6341555 doi: 10.1155/2022/6341555
|
| [71] |
Ajiboye TT, Ayangbenro AS, Babalola OO (2022) Functional Diversity of Microbial Communities in the Soybean (Glycine max L.) Rhizosphere from Free State, South Africa. Int J Mol Sci 23: 9422. https://doi.org/10.3390/ijms23169422 doi: 10.3390/ijms23169422
|
| [72] |
Ayiti OE, Ayangbenro AS, Babalola OO (2022) 16S Amplicon Sequencing of Nitrifying Bacteria and Archaea Inhabiting Maize Rhizosphere and the Influencing Environmental Factors. Agriculture 12: 1328. https://doi.org/10.3390/agriculture12091328 doi: 10.3390/agriculture12091328
|
| [73] |
Adeleke BS, Ayangbenro AS, Babalola OO (2021) Bacterial community structure of the sunflower (Helianthus annuus) endosphere. Plant Signaling Behav 16: 1974217. https://doi.org/10.1080/15592324.2021.1974217 doi: 10.1080/15592324.2021.1974217
|
| [74] |
Simonin M, Dasilva C, Terzi V, et al. (2020) Influence of plant genotype and soil on the wheat rhizosphere microbiome: evidences for a core microbiome across eight African and European soils. FEMS Microbiol Ecol 96: fiaa067. https://doi.org/10.1093/femsec/fiaa067 doi: 10.1093/femsec/fiaa067
|
| [75] |
Mulatu A, Megersa N, Abena T, et al. (2022) Biodiversity of the Genus Trichoderma in the Rhizosphere of Coffee (Coffea arabica) Plants in Ethiopia and Their Potential Use in Biocontrol of Coffee Wilt Disease. Crops 2: 120–141. https://doi.org/10.3390/crops2020010 doi: 10.3390/crops2020010
|
| [76] |
Kaushal M, Tumuhairwe JB, Kaingo J, et al. (2022) Compositional Shifts in Microbial Diversity under Traditional Banana Cropping Systems of Sub-Saharan Africa. Biology 11: 756. https://doi.org/10.3390/biology11050756 doi: 10.3390/biology11050756
|
| [77] |
Abera S, Shimels M, Tessema T, et al. (2022) Back to the roots: defining the core microbiome of Sorghum bicolor in agricultural field soils from the centre of origin. FEMS Microbiol Ecol 98: fiac136. https://doi.org/10.1093/femsec/fiac136 doi: 10.1093/femsec/fiac136
|
| [78] |
Omomowo IO, Adedayo AA, Omomowo OI (2020) Biocontrol Potential of Rhizospheric Fungi from Moringa oleifera, their Phytochemicals and Secondary Metabolite Assessment Against Spoilage Fungi of Sweet Orange (Citrus sinensis). Asian J Appl Sci 8: 1–14. https://doi.org/10.24203/ajas.v8i1.6047 doi: 10.24203/ajas.v8i1.6047
|
| [79] |
Bernardo B, Candeias C, Rocha F (2023) The Contribution of the Hulene-B Waste Dump (Maputo, Mozambique) to the Contamination of Rhizosphere Soils, Edible Plants, Stream Waters, and Groundwaters. Environments 10: 45. https://doi.org/10.3390/environments10030045 doi: 10.3390/environments10030045
|
| [80] |
Sulaiman A, Huan P, Shi-ming LIU, et al. (2022) Molecular and morphological characterization of stunt nematodes of wheat, maize, and rice in the savannahs of northern Nigeria. J Integr Agric 21: 586–595. https://doi.org/10.1016/S2095-3119(21)63696-1 doi: 10.1016/S2095-3119(21)63696-1
|
| [81] |
Owusu Danquah E, Frimpong F, Yeboah S, et al. (2022) Pigeonpea (Cajanus cajan) and white yam (Dioscorea rotundata) cropping system: Improved resource use and productivity in Ghana. Ann Agric Sci 67: 60–71. https://doi.org/10.1016/j.aoas.2022.05.001 doi: 10.1016/j.aoas.2022.05.001
|
| [82] |
Aßhauer KP, Wemheuer B, Daniel R, et al. (2015) Tax4Fun: predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 31: 2882–2884. https://doi.org/10.1093/bioinformatics/btv287 doi: 10.1093/bioinformatics/btv287
|
| [83] |
Du J, Gao Q, Ji C, et al. (2022) Bacillus licheniformis JF-22 to Control Meloidogyne incognita and Its Effect on Tomato Rhizosphere Microbial Community. Front Microbiol 13: 863341. https://doi.org/10.3389/fmicb.2022.863341 doi: 10.3389/fmicb.2022.863341
|
| [84] |
Guerrieri MC, Fanfoni E, Fiorini A, et al. (2020) Isolation and Screening of Extracellular PGPR from the Rhizosphere of Tomato Plants after Long-Term Reduced Tillage and Cover Crops. Plants 9: 668. https://doi.org/10.3390/plants9050668 doi: 10.3390/plants9050668
|
| [85] |
Langille MGI, Zaneveld J, Caporaso JG, et al. (2013) Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat Biotechnol 31: 814–821. https://doi.org/10.1038/nbt.2676 doi: 10.1038/nbt.2676
|
| [86] |
DeSantis TZ, Hugenholtz P, Larsen N, et al. (2006) Greengenes, a Chimera-Checked 16S rRNA Gene Database and Workbench Compatible with ARB. Appl Environ Microbiol 72: 5069–5072. https://doi.org/10.1128/AEM.03006-05 doi: 10.1128/AEM.03006-05
|
| [87] |
Smulders L, Benítez E, Moreno B, et al. (2021) Tomato Domestication Affects Potential Functional Molecular Pathways of Root-Associated Soil Bacteria. Plants 10: 1942. https://doi.org/10.3390/plants10091942 doi: 10.3390/plants10091942
|
| [88] |
Hur M, Park SJ (2019) Identification of Microbial Profiles in Heavy-Metal-Contaminated Soil from Full-Length 16S rRNA Reads Sequenced by a PacBio System. Microorganisms 7: 357. https://doi.org/10.3390/microorganisms7090357 doi: 10.3390/microorganisms7090357
|
| [89] |
Toole DR, Zhao J, Martens-Habbena W, et al. (2021) Bacterial functional prediction tools detect but underestimate metabolic diversity compared to shotgun metagenomics in southwest Florida soils. Appl Soil Ecol 168: 104129. https://doi.org/10.1016/j.apsoil.2021.104129 doi: 10.1016/j.apsoil.2021.104129
|
| [90] |
Babalola OO, Adedayo AA, Fadiji AE (2022) Metagenomic Survey of Tomato Rhizosphere Microbiome Using the Shotgun Approach. Microbiol Resour Announc 11: e01131–01121. https://doi.org/10.1128/mra.01131-21 doi: 10.1128/mra.01131-21
|
| [91] |
Heo AY, Koo YM, Choi HW (2022) Biological Control Activity of Plant Growth Promoting Rhizobacteria Burkholderia contaminans AY001 against Tomato Fusarium Wilt and Bacterial Speck Diseases. Biology 11: 619. https://doi.org/10.3390/biology11040619 doi: 10.3390/biology11040619
|
| [92] |
Gashash EA, Osman NA, Alsahli AA, et al. (2022) Effects of Plant-Growth-Promoting Rhizobacteria (PGPR) and Cyanobacteria on Botanical Characteristics of Tomato (Solanum lycopersicon L.) Plants. Plants 11: 2732. https://doi.org/10.3390/plants11202732 doi: 10.3390/plants11202732
|
| [93] |
Devi R, Kaur T, Kour D, et al. (2020) Beneficial fungal communities from different habitats and their roles in plant growth promotion and soil health. Microbial Biosystems 5: 21–47. https://doi.org/10.21608/mb.2020.32802.1016 doi: 10.21608/mb.2020.32802.1016
|
| [94] | Arya R (2022) Isolation and screening of some rhizosphere fungi for their antagonistic potential against Fusarium oxysporum f. sp. lycopersici in tomato cv Pusa Ruby. Indian J Appl Pure Bio 37: 765–769. |
| [95] |
Roussis I, Beslemes D, Kosma C, et al. (2022) The Influence of Arbuscular Mycorrhizal Fungus Rhizophagus irregularis on the Growth and Quality of Processing Tomato (Lycopersicon esculentum Mill.) Seedlings. Sustainability 14: 9001. https://doi.org/10.3390/su14159001 doi: 10.3390/su14159001
|
| [96] |
Rehan M, Al-Turki A, Abdelmageed AHA, et al. (2023) Performance of Plant-Growth-Promoting Rhizobacteria (PGPR) Isolated from Sandy Soil on Growth of Tomato (Solanum & nbsp; lycopersicum L.). Plants 12: 1588. https://doi.org/10.3390/plants12081588 doi: 10.3390/plants12081588
|
| [97] |
Djebaili R, Pellegrini M, Smati M, et al. (2020) Actinomycete Strains Isolated from Saline Soils: Plant-Growth-Promoting Traits and Inoculation Effects on Solanum lycopersicum. Sustainability 12: 4617. https://doi.org/10.3390/su12114617 doi: 10.3390/su12114617
|
| [98] |
Djebaili R, Pellegrini M, Ercole C, et al. (2021) Biocontrol of Soil-Borne Pathogens of Solanum lycopersicum L. and Daucus carota L. by Plant Growth-Promoting Actinomycetes: In Vitro and In Planta Antagonistic Activity. Pathogens 10: 1305. https://doi.org/10.3390/pathogens10101305 doi: 10.3390/pathogens10101305
|
| [99] |
Kalozoumis P, Savvas D, Aliferis K, et al. (2021) Impact of Plant Growth-Promoting Rhizobacteria Inoculation and Grafting on Tolerance of Tomato to Combined Water and Nutrient Stress Assessed via Metabolomics Analysis. Front Plant Sci 12: 1–15. https://doi.org/10.3389/fpls.2021.670236 doi: 10.3389/fpls.2021.670236
|
| [100] |
Koza NA, Adedayo AA, Babalola OO, et al. (2022) Microorganisms in Plant Growth and Development: Roles in Abiotic Stress Tolerance and Secondary Metabolites Secretion. Microorganisms 10: 1528. https://doi.org/10.3390/microorganisms10081528 doi: 10.3390/microorganisms10081528
|
| [101] |
Adedayo AA, Babalola OO (2023) Fungi That Promote Plant Growth in the Rhizosphere Boost Crop Growth. Journal of Fungi 9: 239. https://doi.org/10.3390/jof9020239 doi: 10.3390/jof9020239
|
| [102] |
Nakahara H, Matsuzoe N, Taniguchi T, et al. (2022) Effect of Burkholderia sp. and Pseudomonas spp. inoculation on growth, yield, and absorption of inorganic components in tomato 'Micro-Tom' under salinity conditions. J Plant Interact 17: 277–289. https://doi.org/10.1080/17429145.2022.2035439 doi: 10.1080/17429145.2022.2035439
|
| [103] |
Zahoor S, Naz R, Keyani R, et al. (2022) Rhizosphere bacteria associated with Chenopodium quinoa promote resistance to Alternaria alternata in tomato. Sci Rep 12: 19027. https://doi.org/10.1038/s41598-022-21857-2 doi: 10.1038/s41598-022-21857-2
|
| [104] |
Dang K, Ji L, Slaughter LC, et al. (2023) Synergistic changes of rhizosphere bacterial community and soil properties in greenhouse soils under long-term tomato monoculture. Appl Soil Ecol 183: 104738. https://doi.org/10.1016/j.apsoil.2022.104738 doi: 10.1016/j.apsoil.2022.104738
|
| [105] |
Rasool M, Akhter A, Soja G, et al. (2021) Role of biochar, compost and plant growth promoting rhizobacteria in the management of tomato early blight disease. Sci Rep 11: 6092. https://doi.org/10.1038/s41598-021-85633-4 doi: 10.1038/s41598-021-85633-4
|
| [106] |
Wang HW, Zhu Y-X, Xu M, et al. (2022) Co-application of spent mushroom substrate and PGPR alleviates tomato continuous cropping obstacle by regulating soil microbial properties. Rhizosphere 23: 100563. https://doi.org/10.1016/j.rhisph.2022.100563 doi: 10.1016/j.rhisph.2022.100563
|
| [107] |
Sarwar MJ, Zahir ZA, Asghar HN, et al. (2023) Co-application of organic amendments and Cd-tolerant rhizobacteria for suppression of cadmium uptake and regulation of antioxidants in tomato. Chemosphere 327: 138478. https://doi.org/10.1016/j.chemosphere.2023.138478 doi: 10.1016/j.chemosphere.2023.138478
|
| [108] |
Devi NO, Tombisana Devi RK, Debbarma M, et al. (2022) Effect of endophytic Bacillus and Arbuscular mycorrhiza fungi (AMF) against Fusarium wilt of tomato caused by Fusarium oxysporum f. sp. lycopersici. Egypt J Biol Pest Control 32. https://doi.org/10.1186/s41938-021-00499-y doi: 10.1186/s41938-021-00499-y
|
| [109] |
Girardi NS, Sosa AL, Etcheverry MG, et al. (2022) In vitro characterization bioassays of the nematophagous fungus Purpureocillium lilacinum: Evaluation on growth, extracellular enzymes, mycotoxins and survival in the surrounding agroecosystem of tomato. Fungal Biol 126: 300–307. https://doi.org/10.1016/j.funbio.2022.02.001 doi: 10.1016/j.funbio.2022.02.001
|
| [110] |
da Silva JM, Dalbon VA, Acevedo JPM, et al. (2022) Development of tomatoes seedlings (Lycopersicum esculentum L.) in combination with silicate rock powder and rhizospheric fungi inoculation. Curr Sci 122: 819. https://doi.org/10.18520/cs/v122/i7/819-825 doi: 10.18520/cs/v122/i7/819-825
|
| [111] |
González-Pérez E, Ortega-Amaro MA, Bautista E, et al. (2022) The entomopathogenic fungus Metarhizium anisopliae enhances Arabidopsis, tomato, and maize plant growth. Plant Physiol Biochem 176: 34–43. https://doi.org/10.1016/j.plaphy.2022.02.008 doi: 10.1016/j.plaphy.2022.02.008
|
| [112] |
De Rocchis V, Jammer A, Camehl I, et al. (2022) Tomato growth promotion by the fungal endophytes Serendipita indica and Serendipita herbamans is associated with sucrose de-novo synthesis in roots and differential local and systemic effects on carbohydrate metabolisms and gene expression. J Plant Physiol 276: 153755. https://doi.org/10.1016/j.jplph.2022.153755 doi: 10.1016/j.jplph.2022.153755
|
| [113] |
Roussis I, Beslemes D, Kosma C, et al. (2022) The Influence of Arbuscular Mycorrhizal Fungus Rhizophagus irregularis on the Growth and Quality of Processing Tomato (Lycopersicon esculentum Mill.) Seedlings. Sustainability 14: 9001. https://doi.org/10.3390/su14159001 doi: 10.3390/su14159001
|
| [114] |
Brooks S, Klomchit A, Chimthai S, et al. (2022) Xylaria feejeensis, SRNE2BP a Fungal Endophyte with Biocontrol Properties to Control Early Blight and Fusarium Wilt Disease in Tomato and Plant Growth Promotion Activity. Curr Microbiol 79: 108. https://doi.org/10.1007/s00284-022-02803-x doi: 10.1007/s00284-022-02803-x
|
| [115] |
Taffner J, Bergna A, Cernava T, et al. (2020) Tomato-Associated Archaea Show a Cultivar-Specific Rhizosphere Effect but an Unspecific Transmission by Seeds. Phytobiomes J 4: 133–141. https://doi.org/10.1094/PBIOMES-01-20-0017-R doi: 10.1094/PBIOMES-01-20-0017-R
|
| [116] |
Li Y, Wang Y, Yu Z, et al. (2018) Effect of inoculum and substrate/inoculum ratio on the performance and methanogenic archaeal community structure in solid state anaerobic co-digestion of tomato residues with dairy manure and corn stover. Waste Manage 81: 117–127. https://doi.org/10.1016/j.wasman.2018.09.042 doi: 10.1016/j.wasman.2018.09.042
|
| [117] |
Osaro-Matthew RC, Ire FS, Frank-Peterside N (2023) Tomato plant growth promotion potential of Janibacter sp strain RC18. Niger J Biotechnol 39: 53–63. https://doi.org/10.4314/njb.v39i2.7 doi: 10.4314/njb.v39i2.7
|
| [118] |
Lahmyed H, Bouharroud R, Qessaoui R, et al. (2021) Actinomycete as Biocontrol Agents against Tomato Gray Mold Disease caused by Botrytis cinerea. Kuwait J Sci 48: 1–8. https://doi.org/10.48129/kjs.v48i3.9200 doi: 10.48129/kjs.v48i3.9200
|
| [119] |
Elshafie HS, Camele I (2022) Rhizospheric Actinomycetes Revealed Antifungal and Plant-Growth-Promoting Activities under Controlled Environment. Plants 11: 1872. https://doi.org/10.3390/plants11141872 doi: 10.3390/plants11141872
|
| [120] |
Singh SP, Gupta R, Gaur R, et al. (2017) Antagonistic Actinomycetes Mediated Resistance in Solanum lycopersicon Mill. against Rhizoctonia solani Kühn. Proceedings of the National Academy of Sciences, India Section B: Biological Sciences 87: 789–798. https://doi.org/10.1007/s40011-015-0651-5 doi: 10.1007/s40011-015-0651-5
|
| [121] |
Nannipieri P, Ascher-Jenull J, Ceccherini MT, et al. (2020) Beyond microbial diversity for predicting soil functions: A mini review. Pedosphere 30: 5–17. https://doi.org/10.1016/S1002-0160(19)60824-6 doi: 10.1016/S1002-0160(19)60824-6
|
| [122] |
Muwawa EM, Obieze CC, Makonde HM, et al. (2021) 16S rRNA Gene Amplicon-based Metagenomic Analysis of Bacterial communities in the Rhizospheres of selected Mangrove species from Mida Creek and Gazi Bay, Kenya. PLoS ONE 16: e0248485. https://doi.org/10.1371/journal.pone.0248485 doi: 10.1371/journal.pone.0248485
|
| [123] |
Omotayo O, Igiehon O, Babalola O (2021) Metagenomic Study of the Community Structure and Functional Potentials in Maize Rhizosphere Microbiome: Elucidation of Mechanisms behind the Improvement in Plants under Normal and Stress Conditions. Sustainability 13: 8079. https://doi.org/10.3390/su13148079 doi: 10.3390/su13148079
|
| [124] |
Dixon M, Simonne E, Obreza T, et al. (2020) Crop Response to Low Phosphorus Bioavailability with a Focus on Tomato. Agron 10: 617. https://doi.org/10.3390/agronomy10050617 doi: 10.3390/agronomy10050617
|
| [125] |
Zhu Z, Bai Y, Lv M, et al. (2020) Soil Fertility, Microbial Biomass, and Microbial Functional Diversity Responses to Four Years Fertilization in an Apple Orchard in North China. Hortic Plant J 6: 223–230. https://doi.org/10.1016/j.hpj.2020.06.003 doi: 10.1016/j.hpj.2020.06.003
|
| [126] |
Chukwuneme CF, Ayangbenro AS, Babalola OO, et al. (2021) Functional diversity of microbial communities in two contrasting maize rhizosphere soils. Rhizosphere 17: 100282. https://doi.org/10.1016/j.rhisph.2020.100282 doi: 10.1016/j.rhisph.2020.100282
|
| [127] |
Xie SY, Ma T, Zhao N, et al. (2022) Whole-Genome Sequencing and Comparative Genome Analysis of Fusarium solani-melongenae Causing Fusarium Root and Stem Rot in Sweetpotatoes. Microbiol Spectrum 10: e00683–00622. https://doi.org/10.1128/spectrum.00683-22 doi: 10.1128/spectrum.00683-22
|
| [128] |
Xin XF, He SY (2013) Pseudomonas syringae pv. tomato DC3000: A Model Pathogen for Probing Disease Susceptibility and Hormone Signaling in Plants. Annu Rev Phytopathol 51: 473–498. https://doi.org/10.1146/annurev-phyto-082712-102321 doi: 10.1146/annurev-phyto-082712-102321
|
| [129] |
Baltrus DA, Nishimura MT, Romanchuk A, et al. (2011) Dynamic Evolution of Pathogenicity Revealed by Sequencing and Comparative Genomics of 19 Pseudomonas syringae Isolates. Pathog 7: e1002132. https://doi.org/10.1371/journal.ppat.1002132 doi: 10.1371/journal.ppat.1002132
|
| [130] |
Stuart JM, Segal E, Koller D, et al. (2003) A Gene-Coexpression Network for Global Discovery of Conserved Genetic Modules. Science 302: 249–255. https://doi.org/10.1126/science.1087447 doi: 10.1126/science.1087447
|
| [131] |
Brüssow H, Canchaya C, Hardt W-D (2004) Phages and the Evolution of Bacterial Pathogens: from Genomic Rearrangements to Lysogenic Conversion. Microbiol Mol Biol Rev 68: 560–602. https://doi.org/10.1128/MMBR.68.3.560-602.2004 doi: 10.1128/MMBR.68.3.560-602.2004
|
| [132] |
Borer ET, Paseka RE, Peace A, et al. (2022) Disease‐mediated nutrient dynamics: Coupling host–pathogen interactions with ecosystem elements and energy. Ecological Monographs 92: 1–22. https://doi.org/10.1002/ecm.1510 doi: 10.1002/ecm.1510
|
| [133] |
Guttman DS, McHardy AC, Schulze-Lefert P (2014) Microbial genome-enabled insights into plant–microorganism interactions. Nat Rev Genet 15: 797–813. https://doi.org/10.1038/nrg3748 doi: 10.1038/nrg3748
|
| [134] |
Bhantana P, Rana MS, Sun X-c, et al. (2021) Arbuscular mycorrhizal fungi and its major role in plant growth, zinc nutrition, phosphorous regulation and phytoremediation. Symbiosis 84: 19–37. https://doi.org/10.1007/s13199-021-00756-6 doi: 10.1007/s13199-021-00756-6
|
| [135] |
Li M, Hou S, Wang J, et al. (2021) Arbuscular mycorrhizal fungus suppresses tomato (Solanum lycopersicum Mill.) Ralstonia wilt via establishing a soil–plant integrated defense system. J Soils Sediments 21: 3607–3619. https://doi.org/10.1007/s11368-021-03016-8 doi: 10.1007/s11368-021-03016-8
|
| [136] |
Yu H, Liu X, Yang C, et al. (2021) Co-symbiosis of arbuscular mycorrhizal fungi (AMF) and diazotrophs promote biological nitrogen fixation in mangrove ecosystems. Soil Biol Biochem 161: 108382. https://doi.org/10.1016/j.soilbio.2021.108382 doi: 10.1016/j.soilbio.2021.108382
|
| [137] |
Yu Y, Gui Y, Li Z, et al. (2022) Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants 11: 386. https://doi.org/10.3390/plants11030386 doi: 10.3390/plants11030386
|
| [138] |
Liu S, Zhang X, Dungait JAJ, et al. (2021) Rare microbial taxa rather than phoD gene abundance determine hotspots of alkaline phosphomonoesterase activity in the karst rhizosphere soil. Biol Fertil Soils 57: 257–268. https://doi.org/10.1007/s00374-020-01522-4 doi: 10.1007/s00374-020-01522-4
|
| [139] |
Sun Q, Hu Y, Chen X, et al. (2021) Flooding and straw returning regulates the partitioning of soil phosphorus fractions and phoD-harboring bacterial community in paddy soils. Appl Microbiol Biotechnol 105: 9343–9357. https://doi.org/10.1007/s00253-021-11672-6 doi: 10.1007/s00253-021-11672-6
|
| [140] |
Wei X, Hu Y, Razavi BS, et al. (2019) Rare taxa of alkaline phosphomonoesterase-harboring microorganisms mediate soil phosphorus mineralization. Soil Biol Biochem 131: 62–70. https://doi.org/10.1016/j.soilbio.2018.12.025 doi: 10.1016/j.soilbio.2018.12.025
|
| [141] |
Xiao H, Yang H, Zhao M, et al. (2021) Soil extracellular enzyme activities and the abundance of nitrogen-cycling functional genes responded more to N addition than P addition in an Inner Mongolian meadow steppe. Sci Total Environ 759: 143541. https://doi.org/10.1016/j.scitotenv.2020.143541 doi: 10.1016/j.scitotenv.2020.143541
|
| [142] |
Zheng MM, Wang C, Li WX, et al. (2021) Changes of acid and alkaline phosphatase activities in long-term chemical fertilization are driven by the similar soil properties and associated microbial community composition in acidic soil. Eur J Soil Biol 104: 103312. https://doi.org/10.1016/j.ejsobi.2021.103312 doi: 10.1016/j.ejsobi.2021.103312
|
| [143] |
Li Y, Wang T, Camps-Arbestain M, et al. (2022) The regulators of soil organic carbon mineralization upon lime and/or phosphate addition vary with depth. Sci Total Environ 828: 154378. https://doi.org/10.1016/j.scitotenv.2022.154378 doi: 10.1016/j.scitotenv.2022.154378
|
| [144] |
Shah A, Nazari M, Antar M, et al. (2021) PGPR in Agriculture: A Sustainable Approach to Increasing Climate Change Resilience. Front Sustain Food Syst 5: 1–22. https://doi.org/10.3389/fsufs.2021.667546 doi: 10.3389/fsufs.2021.667546
|
| [145] |
Xu G, Zhang L, Liu X, et al. (2022) Combined assembly of long and short sequencing reads improve the efficiency of exploring the soil metagenome. BMC Genom 23: 37. https://doi.org/10.1186/s12864-021-08260-3 doi: 10.1186/s12864-021-08260-3
|
| [146] |
Cheng YT, Zhang L, He SY (2019) Plant-Microbe Interactions Facing Environmental Challenge. Cell Host Microbe 26: 183–192. https://doi.org/10.1016/j.chom.2019.07.009 doi: 10.1016/j.chom.2019.07.009
|
| [147] |
Tosi M, Mitter EK, Gaiero J, et al. (2020) It takes three to tango: the importance of Microbes, Host plant, and Soil Management to elucidate manipulation strategies for the Plant Microbiome. Can J Microbiol 66: 413–433. https://doi.org/10.1139/cjm-2020-0085 doi: 10.1139/cjm-2020-0085
|
| [148] |
Dellagi A, Quillere I, Hirel B (2020) Beneficial soil-borne bacteria and fungi: a promising way to improve plant nitrogen acquisition. J Exp Bot 71: 4469–4479. https://doi.org/10.1093/jxb/eraa112 doi: 10.1093/jxb/eraa112
|
| [149] |
Cordovez V, Dini-Andreote F, Carrión VJ, et al. (2019) Ecology and Evolution of Plant Microbiomes. Annu Rev Microbiol 73: 69–88. https://doi.org/10.1146/annurev-micro-090817-062524 doi: 10.1146/annurev-micro-090817-062524
|
| [150] |
Wille L, Messmer MM, Studer B, et al. (2019) Insights to Plant-Microbe interactions provide opportunities to improve Resistance Breeding against Root Diseases in Grain Legumes. Plant Cell Environ 42: 20–40. https://doi.org/10.1111/pce.13214 doi: 10.1111/pce.13214
|


Figures(3) / Tables(2)

Afeez Adesina Adedayo, Samir Ben Romdhane, Olubukola Oluranti Babalola. Metagenomic study of the tomato rhizosphere soil microbiome: current state and prospect[J]. AIMS Geosciences, 2023, 9(2): 330-356. doi: 10.3934/geosci.2023018







 DownLoad:
DownLoad: