Citation: Theresa Vertigan, Kriya Dunlap, Arleigh Reynolds, Lawrence Duffy. Effects of Methylmercury exposure in 3T3-L1 Adipocytes[J]. AIMS Environmental Science, 2017, 4(1): 94-111. doi: 10.3934/environsci.2017.1.94

| [1] | Arctic Monitoring and Assessment Programme (2011) AMAP Assessment 2011: Mercury in the Arctic. Arctic Pollution 2011. |
| [2] |
Krabbenhoft DP, Sunderland EM (2013) Global change and mercury. Science 341: 1457-1458. doi: 10.1126/science.1242838

|
| [3] |
Fisher JA, Jacob DJ, Soerensen AL, et al. (2012) Riverine source of Arctic Ocean mercury inferred from atmospheric observations. Nat Geosci 5: 499-504. doi: 10.1038/ngeo1478
|
| [4] |
Stern GA, Macdonald RW, Outridge PM, et al. (2012). How does climate change influence Arctic mercury? Sci Total Environ 414: 22-42. doi: 10.1016/j.scitotenv.2011.10.039
|
| [5] | Loring PA, Duffy LK (2011) Managing environmental risks: the benefits of a place-based approach. Rural and Remote Health 11: 1800-1808. |
| [6] | Lemire M, Kwan M, Laouan-Sidi AE, et al. (2015) Local country food sources of methylmercury, selenium and omega-3 fatty acids in Nunavik, Northern Quebec. Sci Total Environ 509: 248-259. |
| [7] |
Mahaffey KR (1999) Methylmercury: A new look at the risks. Public Health Reports 114: 397-413. doi: 10.1093/phr/114.5.397
|
| [8] |
Kanda H, Shinkai Y, Kumagai Y (2014) S -Mercuration of cellular proteins by methylmercury and its toxicological implications. J Toxicol Sci 39: 687-700. doi: 10.2131/jts.39.687
|
| [9] |
Weiss B (2007) Why Methylmercury Remains a Conundrum 50 Years after Minamata. Toxicol Sci 97: 223-225. doi: 10.1093/toxsci/kfm047
|
| [10] | Kerper LE, Ballatori N, Clarkson TW (1992) Methylmercury transport across the blood-brain barrier by an amino acid carrier. Am J Physiol-Reg I 262: 761-765. |
| [11] | Kumagai Y, Kanda H, Shinkai Y, et al. (2013) The Role of the Keap1/Nrf2 Pathway in the Cellular Response to Methylmercury. Oxid Med Cell Longev 2013. |
| [12] |
Hirooka T, Yamamoto C, Yasutake A, et al. (2013) Expression of VEGF-related proteins in cultured human brain microvascular endothelial cells and pericytes after exposure to methylmercury. J Toxicol Sci 38: 837-845. doi: 10.2131/jts.38.837
|
| [13] |
He K, Xun P, Liu K, et al. (2013) Mercury exposure in young adulthood and incidence of diabetes later in life: the CARDIA Trace Element Study. Diabetes Care 36: 1584-1589. doi: 10.2337/dc12-1842
|
| [14] |
Futatsuka M, Kitano T, Wakamiya J (1996) An epidemiological study on diabetes mellitus in the population living in a methyl mercury polluted area. J Epidemiol 6: 204-208. doi: 10.2188/jea.6.204
|
| [15] | Yamamoto M, Yanagisawa R, Motomura E, et al. (2013) Increased methylmercury toxicity related to obesity in diabetic KK-Ay mice. J Appl Toxicol 34: 914-923. |
| [16] |
Barnes DM, Hanlon PR, Kircher EA (2003) Effects of inorganic HgCl2 on adipogenesis. Toxicol Sci 75: 368-377. doi: 10.1093/toxsci/kfg195
|
| [17] |
Barnes DM, Kircher EA (2005) Effects of mercuric chloride on glucose transport in 3T3-L1 adipocytes. Toxicol in Vitro 19: 207-214. doi: 10.1016/j.tiv.2004.08.005
|
| [18] |
Cao Y (2013) Angiogenesis and vascular functions in modulation of obesity, adipose metabolism, and insulin sensitivity. Cell Metab 18: 478-489. doi: 10.1016/j.cmet.2013.08.008
|
| [19] |
Elias I, Franckhauser S, Bosch F (2013) New insights into adipose tissue VEGF-A actions in the control of obesity and insulin resistance. Adipocyte 2: 109-112. doi: 10.4161/adip.22880
|
| [20] |
Sun K, Asterholm IW, Kusminski CM, et al. (2012) Dichotomous effects of VEGF-A on adipose tissue dysfunction. Proc Natl Acad Sci USA 109: 5874-5879. doi: 10.1073/pnas.1200447109
|
| [21] | Trayhurn P, Bing C, Wood IS (2005) Adipose Tissue and Adipokines-Energy Regulation from the Inside Out. In The WALTHAM International Nutritional Sciences Symposium (pp. 1935–1939). |
| [22] |
Williams MG (1997) Development of Insulin Resistance in 3T3-L1 Adipocytes. J Biol Chem 272: 7759-7764. doi: 10.1074/jbc.272.12.7759
|
| [23] |
Martini CN, Brandani JN, Gabrielli M, et al. (2014) Effect of hexavalent chromium on proliferation and differentiation to adipocytes of 3T3-L1 fibroblasts. Toxicol in Vitro 28: 700-706. doi: 10.1016/j.tiv.2014.02.003
|
| [24] | American Type 25 Culture Collection (2011) Chemically-Induced Differentiation of ATCC CL-173 (3T3-L1) Using Single-component Commercially-available Reagents. Retrieved February 10, 2015, available from: http://www.atcc.org/~/media/6124AF1E4C2A47CF904435117909AC25.ashx |
| [25] | Hamade AK (2014) Fish Consumption Advice for Alaskans: A Risk Management Strategy to Optimize the Public's Health (pp. 1–78). Section of Epidemiology Division of Public Health Department of Health and Social Services State of Alaska |
| [26] |
Elias I, Franckhauser S, Bosch F (2013) New insights into adipose tissue VEGF-A actions in the control of obesity and insulin resistance. Adipocyte 2: 109-112. doi: 10.4161/adip.22880
|
| [27] |
Sun K, Asterholm IW, Kusminski CM, et al. (2012) Dichotomous effects of VEGF-A on adipose tissue dysfunction. Proc Natl Acad Sci USA 109: 5874-5879. doi: 10.1073/pnas.1200447109
|
| [28] | Gregoire FM, Smas CM, Sul HS (1998) Understanding Adipocyte Differentiation. Physiol Rev 78: 783-810. |
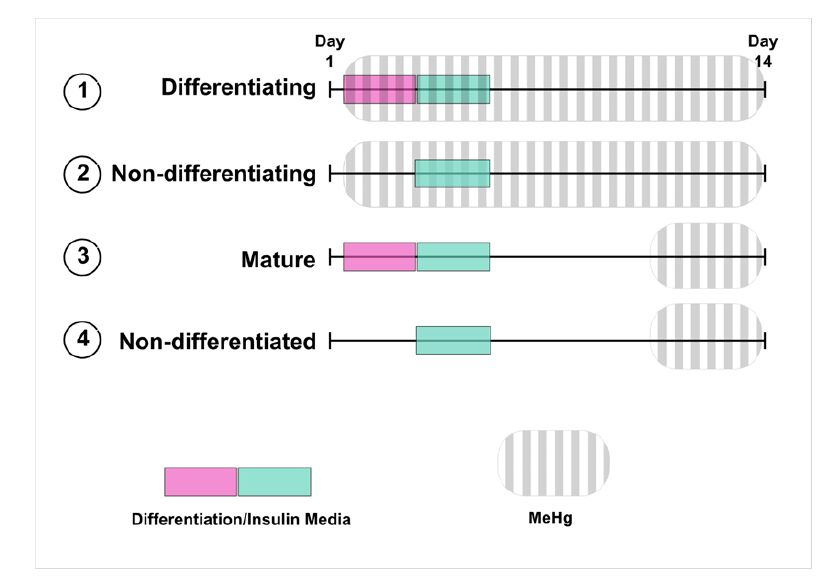

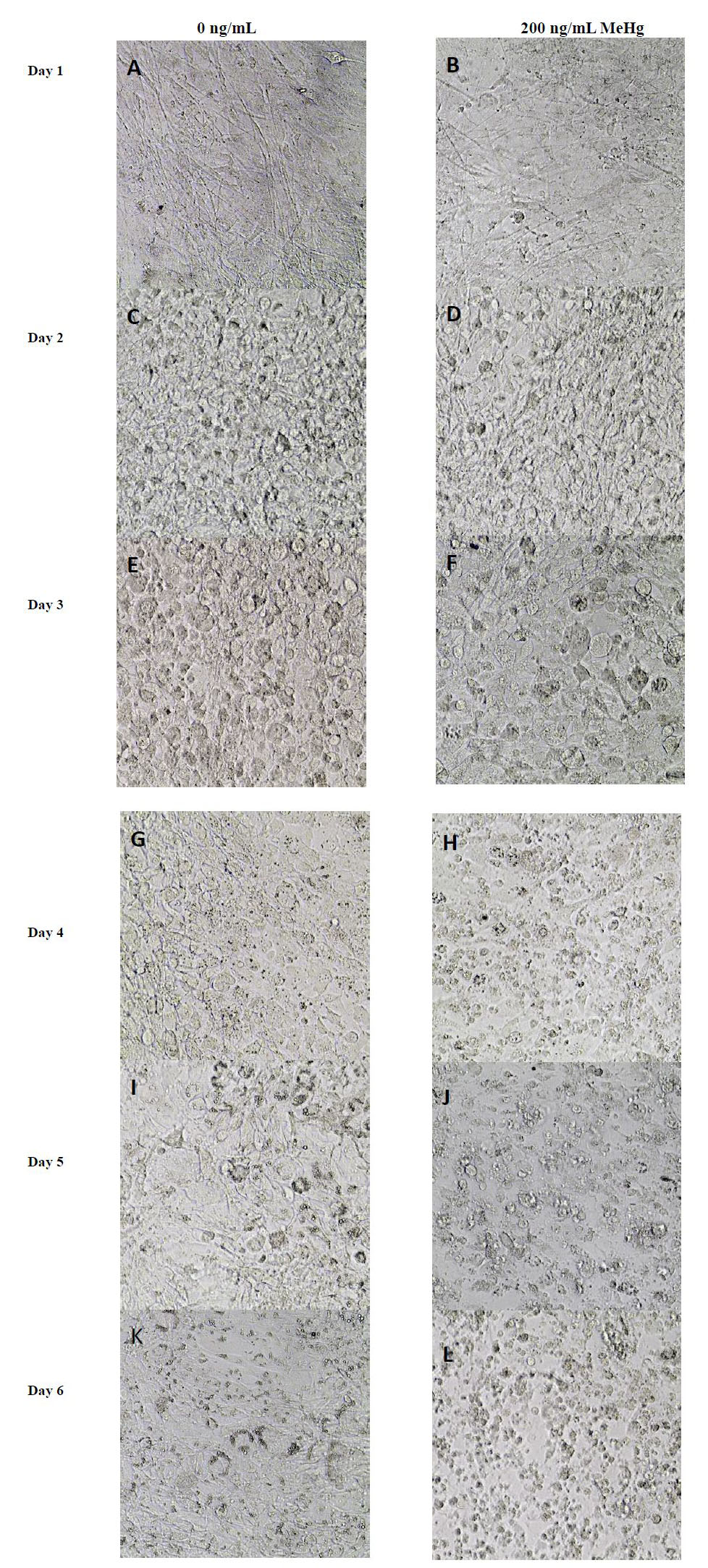
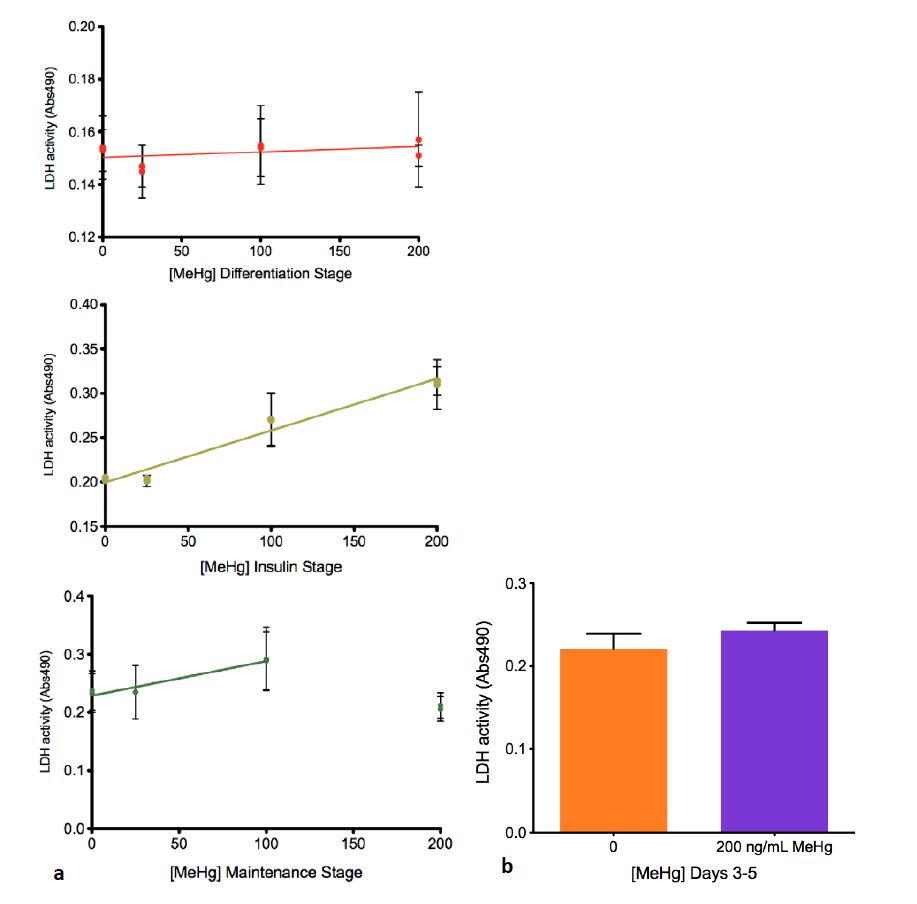
Figures(6) / Tables(1)

Theresa Vertigan, Kriya Dunlap, Arleigh Reynolds, Lawrence Duffy. Effects of Methylmercury exposure in 3T3-L1 Adipocytes[J]. AIMS Environmental Science, 2017, 4(1): 94-111. doi: 10.3934/environsci.2017.1.94







 DownLoad:
DownLoad: