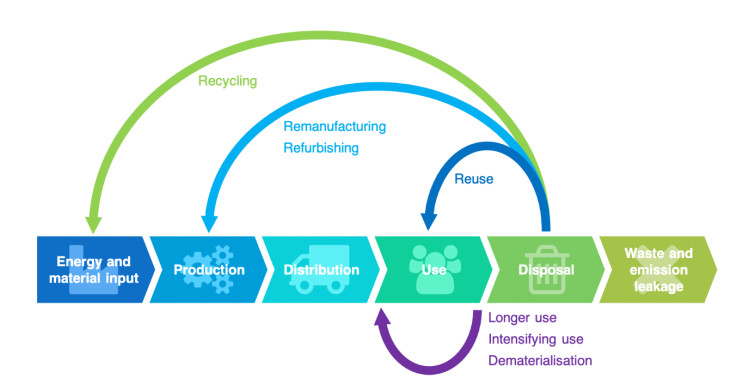
The Paris Agreement goals require a rapid and deep reduction in global greenhouse gas emissions. Recent studies have shown the large potential of circular economy to reduce global emissions by improving resource and material efficiency practices. However, most large-scale energy system and Integrated Assessment Models used for mitigation analysis typically ignore or do not adequately represent circular economy measures. This study aims to fill in this research gap by enhancing a leading global energy system model with a representation of energy efficiency and circular economy considerations. The scenario-based analysis offers an improved understanding of the potentials, costs and impacts of circular economy in the decarbonisation context. The study shows that enhanced energy efficiency and increased material circularity can reduce energy consumption in all sectors, but most importantly in the industrial sector. They can also reduce the required carbon price to achieve Paris goals and the dependence on expensive, immature, and risky technologies, like Carbon Capture and Storage. Circular economy measures should be properly integrated with broad climate policies to provide a holistic and self-consistent framework to deeply reduce carbon emissions.
Citation: Panagiotis Fragkos. Analysing the systemic implications of energy efficiency and circular economy strategies in the decarbonisation context[J]. AIMS Energy, 2022, 10(2): 191-218. doi: 10.3934/energy.2022011
The Paris Agreement goals require a rapid and deep reduction in global greenhouse gas emissions. Recent studies have shown the large potential of circular economy to reduce global emissions by improving resource and material efficiency practices. However, most large-scale energy system and Integrated Assessment Models used for mitigation analysis typically ignore or do not adequately represent circular economy measures. This study aims to fill in this research gap by enhancing a leading global energy system model with a representation of energy efficiency and circular economy considerations. The scenario-based analysis offers an improved understanding of the potentials, costs and impacts of circular economy in the decarbonisation context. The study shows that enhanced energy efficiency and increased material circularity can reduce energy consumption in all sectors, but most importantly in the industrial sector. They can also reduce the required carbon price to achieve Paris goals and the dependence on expensive, immature, and risky technologies, like Carbon Capture and Storage. Circular economy measures should be properly integrated with broad climate policies to provide a holistic and self-consistent framework to deeply reduce carbon emissions.

| [1] | IRP, Global Resources Outlook (2019): Natural Resources for the Future We Want. A Report of the International Resource Panel. 2019, United Nations Environment Programme.: Nairobi, Kenya. Available from: https://www.resourcepanel.org/reports/global-resources-outlook. |
| [2] |
Van der Voet E, Van Oers L, Verboon M, et al. (2018) Environmental implications of future demand scenarios for metals: Methodology and application to the case of seven major metals. J Ind Ecol 23: 141-155. https://doi.org/10.1111/jiec.12722 doi: 10.1111/jiec.12722

|
| [3] |
Hertwich EG (2021) Increased carbon footprint of materials production driven by rise in investments. Nat Geosci 14: 151-155. https://doi.org/10.1038/s41561-021-00690-8 doi: 10.1038/s41561-021-00690-8
|
| [4] |
Rogelj J, Luderer G, Pietzcker RC, et al. (2015) Energy system transformations for limiting end-of-century warming to below 1.5 ℃. Nat Clim Change 5: 519-527. https://doi.org/10.1038/nclimate2572 doi: 10.1038/nclimate2572
|
| [5] |
Kirchherr J, Reike D, Hekkert M (2017) Conceptualizing the circular economy: An analysis of 114 definitions. Resour, Conserv Recycl 127: 221-232. https://doi.org/10.1016/j.resconrec.2017.09.005 doi: 10.1016/j.resconrec.2017.09.005
|
| [6] | Bocken N, Miller K, Evans S (2016) Assessing the environmental impact of new Circular business models. Conference " New Business Models" —Exploring a changing view on organizing value creation—Toulouse, France. Available from: https://www.researchgate.net/publication/305264490_Assessing_the_environmental_impact_of_new_Circular_business_models. |
| [7] | European Commission, Circular Economy Action Plan (2019) Available from: https://ec.europa.eu/environment/strategy/circular-economy-action-plan_en. |
| [8] | European Commission (2018) In-Depth Analysis in Support of the Commission Communication COM (2018) 773, A Clean Planet for all: A European long-term strategic vision for a prosperous, modern, competitive and climate neutral economy. Available from: https://ec.europa.eu/clima/system/files/2018-11/com_2018_733_analysis_in_support_en.pdf. |
| [9] | Material Economics AB (2018) The Circular Economy. Available from: https://materialeconomics.com/publications/the-circular-economy-a-powerful-force-for-climate-mitigation-1. |
| [10] |
Fragkos P, Fragkiadakis K, Paroussos L, et al. (2018) Coupling national and global models to explore policy impacts of NDCs. Energy Policy 118: 462-473. https://doi.org/10.1016/j.enpol.2018.04.002 doi: 10.1016/j.enpol.2018.04.002
|
| [11] |
Pauliuk S, Heeren N, Berrill P, et al. (2021) Global scenarios of resource and emission savings from material efficiency in residential buildings and cars. Nat Commun 12: 5097. https://doi.org/10.1038/s41467-021-25300-4 doi: 10.1038/s41467-021-25300-4
|
| [12] |
Edelenbosch OY, Kermeli K, Crijns-Graus W, et al. (2017) Comparing projections of industrial energy demand and greenhouse gas emissions in long-term energy models. Energy 122: 701-710. https://doi.org/10.1016/j.energy.2017.01.017 doi: 10.1016/j.energy.2017.01.017
|
| [13] |
Geissdoerfer M, Pieroni M, Pigosso D, et al. (2020) Circular Business Models: A Review. J Cleaner Prod 277: 123741. https://doi.org/10.1016/j.jclepro.2020.123741 doi: 10.1016/j.jclepro.2020.123741
|
| [14] | European Commission, Study on the review of the list of critical raw materials: Non-critical raw materials factsheets (2020) Available from: https://op.europa.eu/en/publication-detail/-/publication/6f1e28a7-98fb-11e7-b92d-01aa75ed71a1/language-en. |
| [15] |
Mayer A, Haas W, Wiedenhofer D, et al. (2019) Measuring progress towards a circular economy: A monitoring framework for Economy-wide material loop closing in the EU28. (2019) J Ind Ecol 23: 62-76. https://doi.org/10.1111/jiec.12809 doi: 10.1111/jiec.12809
|
| [16] | UNEP (2017) Resource Efficiency: Potential and Economic Implications. Available from: http://www.resourcepanel.org/reports/resource-efficiency. |
| [17] | European Commission (2018) A European strategy for plastics in a circular economy. Available from: https://www.europarc.org/wp-content/uploads/2018/01/Eu-plastics-strategy-brochure.pdf. |
| [18] | Trinomics (2018) Cooperation fostering industrial symbiosis: market potential, good practice and policy actions. Available from: https://op.europa.eu/en/publication-detail/-/publication/174996c9-3947-11e8-b5fe-01aa75ed71a1/language-en. |
| [19] |
Fragkos P (2020) Global energy system transformations to 1.5 ℃: The impact of revised intergovernmental panel on climate change carbon budgets. Energy Technol 8: 2000395. https://doi.org/10.1002/ente.202000395 doi: 10.1002/ente.202000395
|
| [20] |
Fragkos P, Kouvaritakis N (2018) Model-based analysis of intended nationally determined contributions and 2 ℃ pathways for major economies. Energy 160: 965-978. https://doi.org/10.1016/j.energy.2018.07.030 doi: 10.1016/j.energy.2018.07.030
|
| [21] | Capros P, DeVita A, Tasios N, et al. (2016) EU Reference Scenario 2016—Energy, Transport and GHG Emissions Trends to 2050; European Commission Directorate General for Energy, Directorate General for Climate Action and Directorate General for Mobility and Transport: Brussels, Belgium, 2016. Available from: http://www.e3mlab.eu/e3mlab/reports/referencescenario2016report.pdf. |
| [22] |
Fragkos P, Kouvaritakis N, Capros P (2015) Incorporating uncertainty into world energy modelling: The Prometheus model. Environ Model Assess 20: 549-569. https://doi.org/10.1007/s10666-015-9442-x doi: 10.1007/s10666-015-9442-x
|
| [23] |
Fragkos P, Kouvaritakis N (2018) Investments in power generation under uncertainty—a MIP specification and Large-Scale application for EU. Environ Model Assess 23: 511-527. https://doi.org/10.1007/s10666-017-9583-1 doi: 10.1007/s10666-017-9583-1
|
| [24] |
Grubler A, Wilson C, Bento N, et al. (2018) A low energy demand scenario for meeting the 1.5 C target and sustainable development goals without negative emission technologies. Nature Energy 3: 515-527. https://doi.org/10.1038/s41560-018-0172-6 doi: 10.1038/s41560-018-0172-6
|
| [25] |
Fotiou T, de Vita A, Capros P (2019) Economic-Engineering modelling of the buildings sector to study the transition towards deep decarbonisation in the EU. Energies 12: 2745. https://doi.org/10.3390/en12142745 doi: 10.3390/en12142745
|
| [26] |
Capros P, Zazias G, Evangelopoulou S, et al. (2019) Energy-system modelling of the EU strategy towards climate-neutrality. Energy Policy 134: 110960. https://doi.org/10.1016/j.enpol.2019.110960 doi: 10.1016/j.enpol.2019.110960
|
| [27] | Towards the Circular Economy vol. 1, 2013, Ellen McArthur Foundation. Available from: https://ellenmacarthurfoundation.org/towards-the-circular-economy-vol-1-an-economic-and-business-rationale-for-an. |
| [28] | IEA (2020) Iron and Steel. Paris. Available from: https://www.iea.org/reports/iron-and-steel. |
| [29] | ISRI (2019) 2019 Recycling Industry Year Book. Available from: https://www.isri.org/recycling-commodities-old/recycling-industry-yearbook. |
| [30] | WSP, Parsons Brinckerhoff, DNV GL (2015) Industrial Decarbonisation & Energy Efficiency Roadmaps to 2050—Cement. Available from: https://www.gov.uk/government/publications/industrial-decarbonisation-and-energy-efficiency-roadmaps-to-2050. |
| [31] | Accenture (2017) Taking the EU chemicals industry into the circular economy. Available from: https://www.accenture.com/us-en/_acnmedia/PDF-45/Accenture-CEFIC-Report-Exec-Summary.pdf. |
| [32] | JRC (2018) Prospective scenarios for the pulp and paper industry. Available from: https://publications.jrc.ec.europa.eu/repository/handle/JRC111652. |
| [33] | van Soest HL, Aleluia Reis L, Baptista LB, et al. (2021) Global roll-out of comprehensive policy measures may aid in bridging emissions gap. Nat Commun, 12. https://doi.org/10.1038/s41467-021-26595-z |
| [34] |
Fragkos P (2021) Assessing the role of carbon capture and storage in mitigation pathways of developing economies. Energies 14: 1879. https://doi.org/10.3390/en14071879 doi: 10.3390/en14071879
|
| [35] |
McCollum DL, Zhou W, Bertram C, et al. (2018) Energy investment needs for fulfilling the Paris Agreement and achieving the Sustainable Development Goals. Nat Energy 3: 589-599. https://doi.org/10.1038/s41560-018-0179-z doi: 10.1038/s41560-018-0179-z
|
| [36] | Rogelj J, Shindell D, Jiang K, et al. (2018) Mitigation pathways compatible with 1.5 ℃ in the context of sustainable development. In Global Warming of 1.5 ℃: an IPCC special report on the impacts of global warming of 1.5 ℃ above pre-industrial levels, Geneva, Switzerland: IPCC, In press. Available from: https://www.ipcc.ch/sr15/. |
| [37] |
Fricko O, Havlik P, Rogelj J, et al. (2017) The marker quantification of the shared socioeconomic pathway 2: A middle-of-the-road scenario for the 21st century. Global Environ Change 42: 251-267. https://doi.org/10.1016/j.gloenvcha.2016.06.004 doi: 10.1016/j.gloenvcha.2016.06.004
|
| [38] | OECD (2020) OECD Economic Outlook; OECD Publishing: Paris, France, 2020; Volume 2020. Available from: https://www.oecd-ilibrary.org/economics/oecd-economic-outlook/volume-2020/issue-1_0d1d1e2e-en#:~:text=GDP%20is%20projected%20to%20fall%20by%2014%25%20in%202020%20before,recover%20by%207.7%25%20in%202021. |
| [39] | The World Bank (2021). Global Economic Prospects (Issue June 2021). Available from: https://www.worldbank.org/en/publication/global-economic-prospects. |
| [40] |
Rochedo PRR, Fragkos P, Garaffa R, et al. (2021) Is green recovery enough? Analysing the impacts of Post-COVID-19 economic packages. Energies 14: 5567. https://doi.org/10.3390/en14175567 doi: 10.3390/en14175567
|
| [41] |
Fragkos PK, Fragkiadakis B, Sovacool L, et al. (2021) Equity implications of climate policy: Assessing the social and distributional impacts of emission reduction targets in the European Union. Energy 237: 21591. https://doi.org/10.1016/j.energy.2021.121591 doi: 10.1016/j.energy.2021.121591
|
| [42] | Vona F (2019) Job losses and political acceptability of climate policies: why the 'job-killing' argument is so persistent and how to overturn it. Climate Policy 19: 524-532. 10.1080/14693062.2018.1532871 |
| [43] |
Madeddu S, Ueckerdt F, Pehl M, et al. (2020) The CO2 reduction potential for the European industry via direct electrification of heat supply (power-to-heat). Environ Res Lett 15: 124004. https://doi.org/10.1088/1748-9326/abbd02 doi: 10.1088/1748-9326/abbd02
|
| [44] |
Levesque A, Pietzker R, Luderer G (2019) Halving energy demand from buildings: The impact of low consumption practices. Technol Forecast Soc Change 146: 253-266. https://doi.org/10.1016/j.techfore.2019.04.025 doi: 10.1016/j.techfore.2019.04.025
|
| [45] |
Rodrigues R, Pietzker R, Fragkos P, et al. (2021) Narrative-driven alternative roads to achieve mid-century CO2 net neutrality in Europe. Energy 239: 121908. https://doi.org/10.1016/j.energy.2021.121908 doi: 10.1016/j.energy.2021.121908
|
| [46] |
Brodny J, Tutak M (2022) Analysis of the efficiency and structure of energy consumption in the industrial sector in the European Union countries between 1995 and 2019. Sci Total Environ 808: 152052. https://doi.org/10.1016/j.scitotenv.2021.152052 doi: 10.1016/j.scitotenv.2021.152052
|
| [47] | Falcone PM, Hiete M, Sapio A (2021) Hydrogen economy and sustainable development goals: Review and policy insights. Curr Opin Green Sustainable Chem 31: 100506. https://doi.org/10.1016/j.cogsc.2021.100506 |
| [48] | Sharma HB, Vanapalli KR, Samal B, et al. (2021) Circular economy approach in solid waste management system to achieve UN-SDGs: Solutions for post-COVID recovery. Sci Total Environ 800, 149605. https://doi.org/10.1016/j.scitotenv.2021.149605 |
| [49] |
Rocca R, Rosa P, Sassanelli C, et al. (2020) Integrating virtual reality and digital twin in circular economy practices: A laboratory application case. Sustainability 12: 2286. https://doi.org/10.3390/su12062286 doi: 10.3390/su12062286
|
| [50] |
D'Adamo I, Falcone PM, Martin M, et al. (2020) A sustainable revolution: Let's go sustainable to get our globe cleaner. Sustainability 12: 4387. https://doi.org/10.3390/su12114387 doi: 10.3390/su12114387
|
| [51] |
Longa FD, Fragkos P, Nogueira LP, et al. (2022) System-level effects of increased energy efficiency in global low-carbon scenarios: A model comparison. Comput Ind Eng 167: 108029. https://doi.org/10.1016/j.cie.2022.108029 doi: 10.1016/j.cie.2022.108029
|
| [52] | Bordage F (2019) The Environmental Footprint of the Digital World. GreenIT.fr, p. 39. Available from: https://www.greenit.fr/wp-content/uploads/2019/11/GREENIT_EENM_etude_EN_accessible.pdf. |










Figures(11) / Tables(3)

Panagiotis Fragkos. Analysing the systemic implications of energy efficiency and circular economy strategies in the decarbonisation context[J]. AIMS Energy, 2022, 10(2): 191-218. doi: 10.3934/energy.2022011







 DownLoad:
DownLoad: