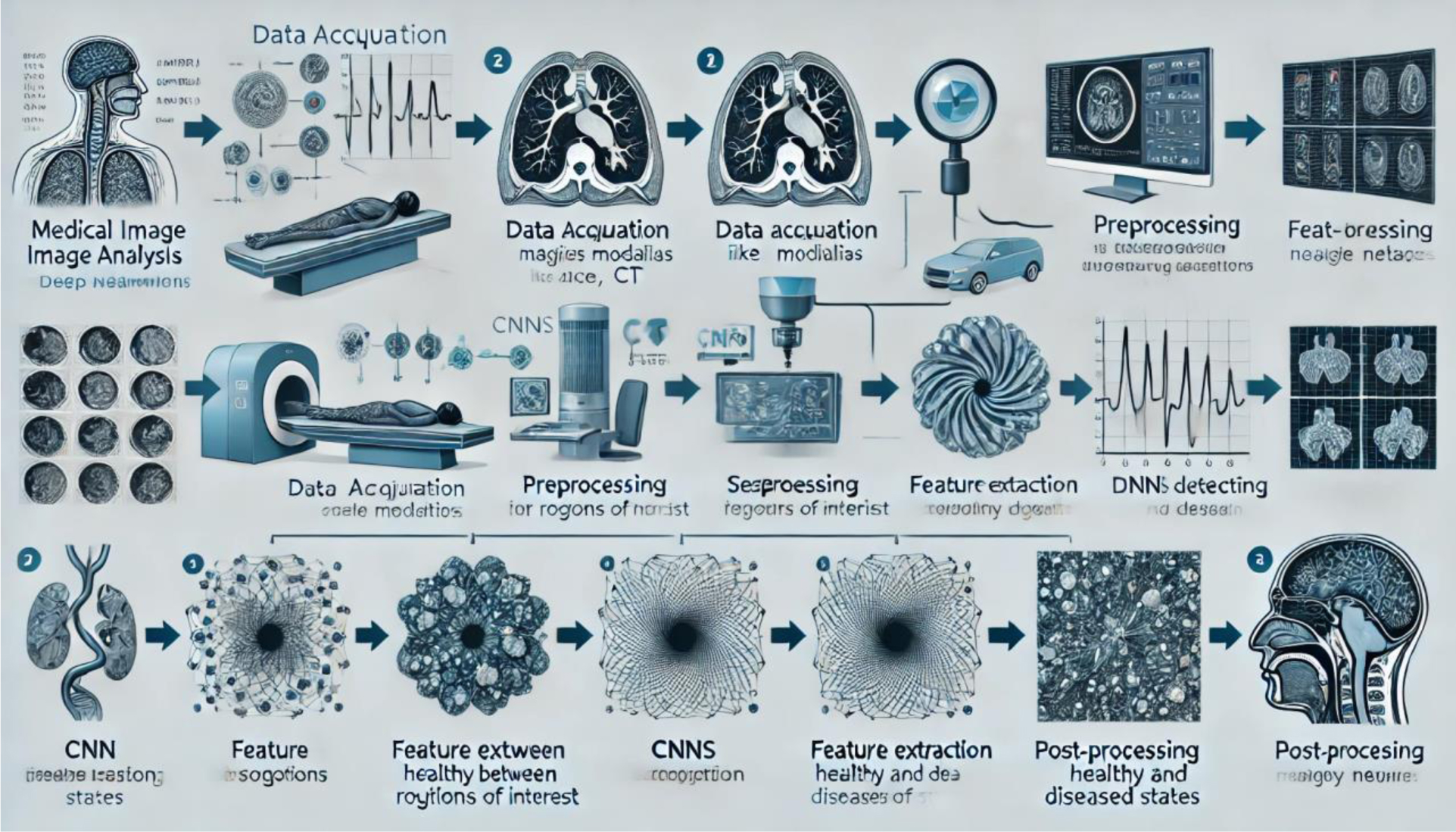
Deep Learning Algorithms (DLAs) have emerged as transformative tools in medical image analysis, offering unprecedented accuracy and efficiency in diagnostic tasks. We explored the state-of-the-art applications of DLAs in medical imaging, focusing on their role in disease detection, segmentation, workflow automation, and multi-modality data integration. Key architectures such as Convolutional Neural Networks (CNNs), U-Net, and Vision Transformers are highlighted, alongside their tailored applications in healthcare. Additionally, Mamba networks have shown significant promise in medical imaging by leveraging their advanced memory-efficient architecture for high-dimensional data processing. These networks excel in real-time analysis, improving the speed and accuracy of complex imaging tasks such as tumor detection and organ segmentation. The adaptability and computational efficiency of Mamba networks position them as a strong alternative to traditional deep learning architectures in the field of medical imaging. DLAs have consistently demonstrated superior performance compared to radiologists in various diagnostic tasks, such as breast cancer detection and brain tumor segmentation, with higher accuracy and efficiency. Despite these advancements, challenges such as limited data availability, ethical concerns, interpretability issues, and integration hurdles persist. Addressing these barriers is crucial to unlocking the full potential of DLAs and enabling their seamless integration into clinical workflows, ultimately enhancing patient care and diagnostic precision.
Citation: Dafina Xhako, Niko Hyka, Elda Spahiu, Suela Hoxhaj. Medical image analysis using deep learning algorithms (DLA)[J]. AIMS Biophysics, 2025, 12(2): 121-143. doi: 10.3934/biophy.2025008
Deep Learning Algorithms (DLAs) have emerged as transformative tools in medical image analysis, offering unprecedented accuracy and efficiency in diagnostic tasks. We explored the state-of-the-art applications of DLAs in medical imaging, focusing on their role in disease detection, segmentation, workflow automation, and multi-modality data integration. Key architectures such as Convolutional Neural Networks (CNNs), U-Net, and Vision Transformers are highlighted, alongside their tailored applications in healthcare. Additionally, Mamba networks have shown significant promise in medical imaging by leveraging their advanced memory-efficient architecture for high-dimensional data processing. These networks excel in real-time analysis, improving the speed and accuracy of complex imaging tasks such as tumor detection and organ segmentation. The adaptability and computational efficiency of Mamba networks position them as a strong alternative to traditional deep learning architectures in the field of medical imaging. DLAs have consistently demonstrated superior performance compared to radiologists in various diagnostic tasks, such as breast cancer detection and brain tumor segmentation, with higher accuracy and efficiency. Despite these advancements, challenges such as limited data availability, ethical concerns, interpretability issues, and integration hurdles persist. Addressing these barriers is crucial to unlocking the full potential of DLAs and enabling their seamless integration into clinical workflows, ultimately enhancing patient care and diagnostic precision.

| [1] | Thakur GK, Thakur A, Kulkarni S, et al. (2024) Deep learning approaches for medical image analysis and diagnosis. Cureus 16: e59507. https://doi.org/10.7759/cureus.59507 |
| [2] | Mienye D, Swart G (2024) A comprehensive review of deep learning: architectures, recent advances, and applications. Information 15: 755. https://doi.org/10.3390/info15120755 |
| [3] | Ronneberger O, Fischer P, Brox T (2015) U-Net: Convolutional networks for biomedical image segmentation. International Conference on Medical Image Computing and Computer-Assisted Intervention . Springer 234-241. https://doi.org/10.1007/978-3-319-24574-4_28 |
| [4] | Zhou Z, Siddiquee R, Tajbakhsh N, et al. (2018) UNet++: A nested U-Net architecture for medical image segmentation. International Conference on Medical Image Computing and Computer-Assisted Intervention . Springer 3-11. https://doi.org/10.1007/978-3-030-00889-5_1 |
| [5] | Xie Y, Zhang Z, Lin H (2019) Deep learning for medical image analysis: a survey. IEEE Access 7: 19064-19079. https://doi.org/10.1109/ACCESS.2019.2899452 |
| [6] |
Tajbakhsh N, Shin Y, Gurudu R, et al. (2020) Convolutional neural networks for medical image analysis: full training or fine-tuning?. IEEE T Med Imaging 35: 1299-1312. https://doi.org/10.1109/TMI.2016.2581847 
|
| [7] |
Cheng Z, Ni D, Chou H, et al. (2016) Computer-aided diagnosis with deep learning: applications in cardiothoracic imaging. J Med Imaging 3: 043502. https://doi.org/10.1117/1.JMI.3.4.043502
|
| [8] |
Li M, Jiang Y, Zhang Y, et al. (2023) Medical image analysis using deep learning algorithms. Front Public Health 11: 1273253. https://doi.org/10.3389/fpubh.2023.1273253
|
| [9] |
Rajpurkar P, Irvin J, Ball RL, et al. (2018) Deep learning for chest radiograph diagnosis: a retrospective comparison of the CheXNeXt algorithm to practicing radiologists. Plos Med 15: e1002686. https://doi.org/10.1371/journal.pmed.1002686
|
| [10] | Nagarjun A, Manju N, Aradhya M, et al. (2024) Classification of skin cancer using deep learning. Proceedings of the 2024 Conference on Skin Cancer Research . Springer 1-10. https://doi.org/10.1007/978-981-99-7633-1_43 |
| [11] |
McKinney SM, Sieniek M, Godbole V, et al. (2020) International evaluation of an AI system for breast cancer screening. Nature 577: 89-94. https://doi.org/10.1038/s41586-019-1799
|
| [12] |
Lundervold AS, Lundervold A (2019) An overview of deep learning in medical imaging focusing on MRI. Z Med Phys 29: 102-127. https://doi.org/10.1016/j.zemedi.2018.11.00
|
| [13] |
Celard P, Iglesias L, Sorribes-Fdez M, et al. (2023) A survey on deep learning applied to medical images: from simple artificial neural networks to generative models. Neural Comput Appl 35: 2291-2323. https://doi.org/10.1007/s00521-022-07953-4
|
| [14] |
Khan I, Rahman A, Debnath T, et al. (2022) Accurate brain tumour detection using deep convolutional neural network. Comput Struct Biotechnol J 20: 4733-4745. https://doi.org/10.1016/j.csbj.2022.08.039
|
| [15] |
Vaz TF, Canto Moreira N, Hellström-Westas L, et al. (2024) Brain extraction methods in neonatal brain MRI and their effects on intracranial volumes. Appl Sci 14: 1339. https://doi.org/10.3390/app14041339
|
| [16] | Xhako D, Spahiu E, Hyka N, et al. (2023) Integration of DCNN model for brain tumour detection with PPIR simulator. Int J Intell Syst Appl Eng 12: 534-538. https://ijisae.org/index.php/IJISAE/article/view/4184 |
| [17] | Xhako D, Hoxhaj S, Hyka N, et al. (2023) Artificial intelligence in medical image processing. Int J Intell Syst Appl Eng 12: 549-552. https://ijisae.org/index.php/IJISAE/article/view/4186 |
| [18] | Xhako D, Spahiu E, Hoxhaj S, et al. (2024) The application of AI-based techniques for early detection of breast cancer (BC). The International Conference on Radiation Applications (RAP 2024) . https://doi.org/10.37392/RapProc.2024.07 |
| [19] | Xhako D, Spahiu E, Hoxhaj S, et al. (2024) Application of AI in higher education: evidence and challenges. Nanotechnol Perceptions 20: S11. https://doi.org/10.62441/nano-ntp.v20iS11.2392 |
| [20] |
Walsh J, Othmani A, Jain M, et al. (2022) Using U-Net network for efficient brain tumor segmentation in MRI images. Healthcare Anal 2: 100098. https://doi.org/10.1016/j.health.2022.100098
|
| [21] | Akcay S, Kundegorski ME, Deena S, et al. (2021) Transfer learning using convolutional neural networks for object classification within X-ray baggage security imagery. Pattern Recogn Lett 105: 171-179. https://doi.org/10.1016/j.patrec.2018.02.005 |
| [22] |
Anthimopoulos M, Christodoulidis S, Ebner L, et al. (2021) Lung pattern classification for interstitial lung diseases using a deep convolutional neural network. IEEE T Med Imaging 35: 1207-1216. https://doi.org/10.1109/TMI.2016.2528229
|
| [23] | Benjamins MR, Yang TC (2022) Machine learning in healthcare disparities: applications and future directions. Health Affair 41: 499-507. https://doi.org/10.1377/hlthaff.2021.01909 |
| [24] | Chen J, Tan C, Hu X (2022) Deep learning for medical image segmentation: advances and applications. Med Phys 49: e215-e234. https://doi.org/10.1002/mp.15320 |
| [25] |
Xia Q, Zheng H, Zou H, et al. (2025) A comprehensive review of deep learning for medical image segmentation. Neurocomputing 613: 128740. https://doi.org/10.1016/j.neucom.2024.128740
|
| [26] |
Kim S, Park J, Son Y, et al. (2025) Development and validation of a machine learning algorithm for predicting diabetes retinopathy in patients with type 2 diabetes: algorithm development study. JMIR Med Inf 13: e58107. https://doi.org/10.2196/58107
|
| [27] |
Shafiq M, Gu Z (2022) Deep residual learning for image recognition: a survey. Appl Sci 12: 8972. https://doi.org/10.3390/app12188972
|
| [28] |
Mall PK, Singh PK, Srivastav S, et al. (2023) A comprehensive review of deep neural networks for medical image processing: recent developments and future opportunities. Healthcare Anal 4: 100216. https://doi.org/10.1016/j.health.2023.100216
|
| [29] |
Hosny A, Parmar C, Quackenbush J, et al. (2018) Artificial intelligence in radiology. Nat Rev Cancer 18: 500-510. https://doi.org/10.1038/s41568-018-0016-5
|
| [30] |
Yuan J (2024) Performance analysis of deep learning algorithms implemented using PyTorch in image recognition. Proc Comput Sci 247: 61-69. https://doi.org/10.1016/j.procs.2024.10.008
|
| [31] |
Litjens G, Kooi T, Bejnordi B, et al. (2017) A survey on deep learning in medical image analysis. Med Image Anal 42: 60-88. https://doi.org/10.1016/j.media.2017.07.005
|
| [32] |
Lundervold AS, Lundervold A (2019) An overview of deep learning in medical imaging focusing on MRI. Z Med Phys 29: 102-127. https://doi.org/10.1016/j.zemedi.2018.11.002
|
| [33] |
Shen D, Wu G, Suk HI (2017) Deep learning in medical image analysis. Annu Rev Biomed Eng 19: 221-248. https://doi.org/10.1146/annurev-bioeng-071516-044442
|
| [34] | Faycel A, Gattal A, Aouine M, et al. (2024) Image classification using a deep convolutional neural network. Proceedings of the 2024 conference on image processing . Springer 1-10. https://doi.org/10.1007/978-3-031-60594-9_5 |
| [35] | Szegedy C, Liu W, Jia Y, et al. (2015) Going deeper with convolutions. Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition : 1-9. https://doi.org/10.1109/CVPR.2015.7298594 |
| [36] |
Wang G, Li W, Aertsen M, et al. (2019) Deep learning in medical ultrasound analysis: a review. Engineering 5: 261-275. https://doi.org/10.1016/j.eng.2018.11.020
|
| [37] |
Yamashita R, Nishio M, Do RKG, et al. (2018) Convolutional neural networks: an overview and application in radiology. Insights Imaging 9: 611-629. https://doi.org/10.1007/s13244-018-0639-9
|
| [38] |
Zhang Z, Jiang W, Maguire L, et al. (2021) Recent advances in medical image analysis using deep learning. Comput Med Imag Grap 89: 101884. https://doi.org/10.1016/j.compmedimag.2021.101884
|
| [39] | Chen W, Liu B, Peng J, et al. (2020) Deep learning for COVID-19 detection and diagnosis from medical imaging. IEEE T Med Imaging 39: 2424-2434. https://doi.org/10.1109/TMI.2020.3003926 |
| [40] |
Singh G, Janghel RR (2022) Deep learning-based frameworks for early detection of Alzheimer's disease. Neural Comput Appl 34: 1635-1650. https://doi.org/10.1007/s00521-021-06188-6
|
| [41] |
Ranjbarzadeh R, Zarbakhsh P (2024) Brain tumor segmentation based on optimized convolutional neural network and improved chimp optimization algorithm. Comput Biol Med 168: 107723. https://doi.org/10.1016/j.compbiomed.2023.107723
|
| [42] | Zhang Y, Tian Z, Kong Y (2018) Feature learning for medical imaging: recent advances and future directions. Neurocomputing 275: 232-240. https://doi.org/10.1016/j.neucom.2017.04.015 |
| [43] |
Yihao L, Mostafa D, Pierre-Henri C, et al. (2024) A review of deep learning-based information fusion techniques for multimodal medical image classification. Comput Biol Med 177: 108635. https://doi.org/10.1016/j.compbiomed.2024.108635
|
| [44] | Maier A, Scheidl J, Hornegger J (2022) Deep learning in computational pathology: trends and advancements. J Pathol Informatics 13: 107-117. https://doi.org/10.4103/jpi.jpi_15_22 |
| [45] | Zeng G, Wang L, Zheng W, et al. (2021) A hybrid deep learning framework for segmentation of liver lesions. Pattern Recogn 123: 108234. https://doi.org/10.1016/j.patcog.2021.108234 |
| [46] | Liu X, Song L, Xu H (2019) Deep reinforcement learning for automated medical image segmentation. Artif Intell Med 101: 101741. https://doi.org/10.1016/j.artmed.2019.101741 |
| [47] | Zhao X, Wu S, Yang Q (2021) GAN-based frameworks for improving medical image synthesis and enhancement. Med Phys 48: 3161-3173. https://doi.org/10.1002/mp.14882 |
| [48] |
Sharma V, Nillmani S, Shukla K (2024) Deep learning models for tuberculosis detection and infected region visualization in chest X-ray images. Intell Med 4: 104-113. https://doi.org/10.1016/j.imed.2023.06.001
|
| [49] | Bansal S, Madisetty S, Rehman MZ, et al. A comprehensive survey of Mamba architectures for medical image analysis: classification, segmentation, restoration, and beyond (2024). https://doi.org/10.48550/arXiv.2410.02362 |
| [50] | Yue Y, Li Z MedMamba: vision Mamba for medical image classification (2024). https://doi.org/10.48550/arXiv.2403.03849 |







Figures(8) / Tables(5)

Dafina Xhako, Niko Hyka, Elda Spahiu, Suela Hoxhaj. Medical image analysis using deep learning algorithms (DLA)[J]. AIMS Biophysics, 2025, 12(2): 121-143. doi: 10.3934/biophy.2025008







 DownLoad:
DownLoad: