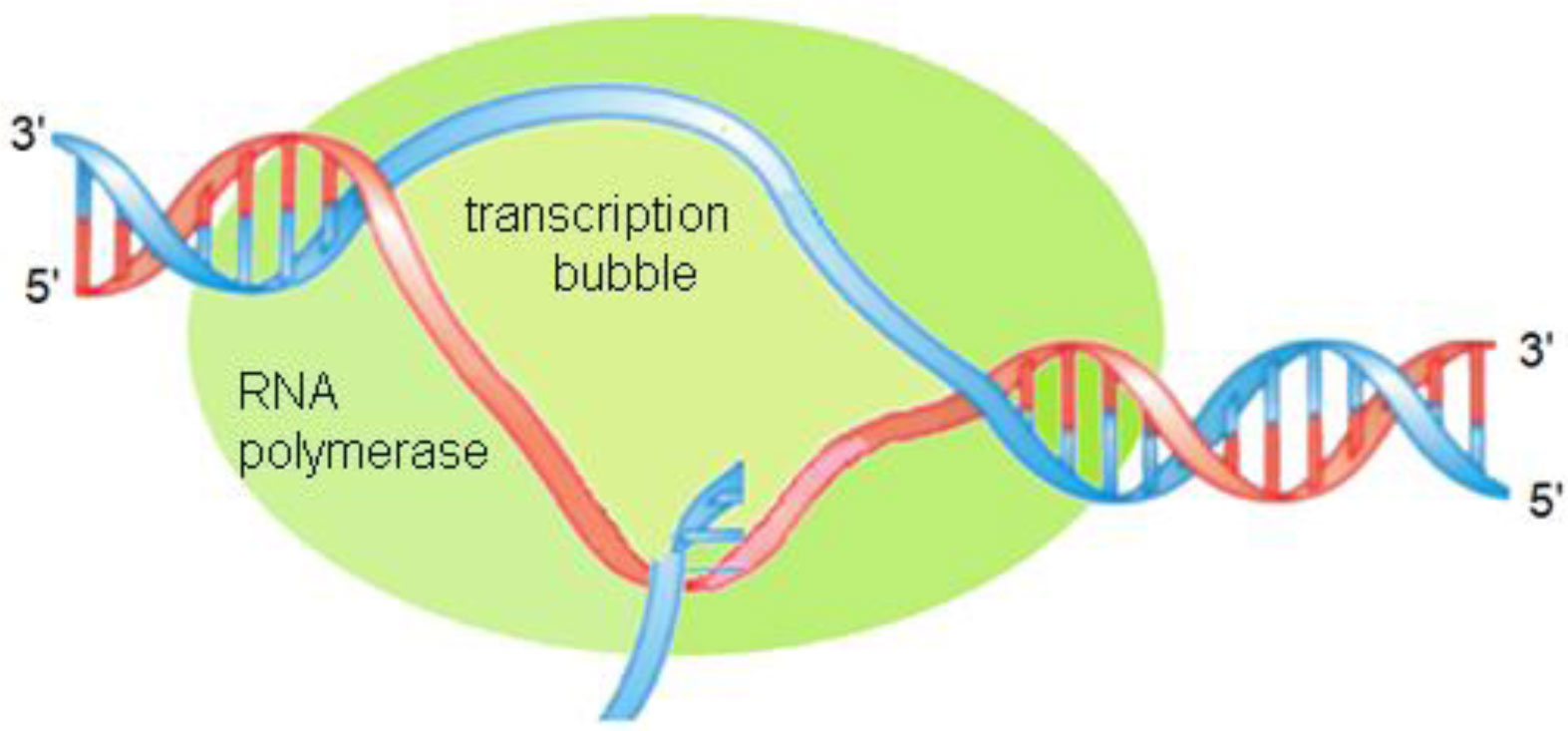
A small circular DNA - plasmid pBR322, is considered as a complex dynamic system where nonlinear conformational perturbations which are often named open states or kinks, can arise and propagate. To describe the internal dynamics of the plasmid we use mathematical model consisting of two coupled sine-Gordon equations that in the average field approximation are transformed to two sine-Gordon independent equations with renormalized coefficients. The first equation describes angular oscillations of nitrous bases of the main chain. The second equation describes angular oscillations of nitrous bases in the complementary chain. As a result, two types of kink-like solutions have been obtained. One type kinks were the solutions of the first equation, and the other kinks were the solutions of the second equation. We calculated the main characteristics of the kink motion, including the time dependences of the kink velocity, coordinates, and total energy. These calculations were performed at the initial velocity equal to 1881 m/s which was chosen to avoid reflections from energy barriers corresponding to CDS-1 and CDS-2. The movement of the kinks was investigated by the method of the double energy profile. The maximum complete set of the DNA dynamic parameters was used to calculate the double profile. To calculate the velocity, energy and trajectory of the kinks, the block method was used. The results obtained made it possible to explain in which region of the plasmid the formation of a transcription bubble is most likely, as well to understand in which direction the bubble will move and the transcription process will go.
Citation: Ludmila V. Yakushevich, Larisa A. Krasnobaeva. Double energy profile of pBR322 plasmid[J]. AIMS Biophysics, 2021, 8(2): 221-232. doi: 10.3934/biophy.2021016
A small circular DNA - plasmid pBR322, is considered as a complex dynamic system where nonlinear conformational perturbations which are often named open states or kinks, can arise and propagate. To describe the internal dynamics of the plasmid we use mathematical model consisting of two coupled sine-Gordon equations that in the average field approximation are transformed to two sine-Gordon independent equations with renormalized coefficients. The first equation describes angular oscillations of nitrous bases of the main chain. The second equation describes angular oscillations of nitrous bases in the complementary chain. As a result, two types of kink-like solutions have been obtained. One type kinks were the solutions of the first equation, and the other kinks were the solutions of the second equation. We calculated the main characteristics of the kink motion, including the time dependences of the kink velocity, coordinates, and total energy. These calculations were performed at the initial velocity equal to 1881 m/s which was chosen to avoid reflections from energy barriers corresponding to CDS-1 and CDS-2. The movement of the kinks was investigated by the method of the double energy profile. The maximum complete set of the DNA dynamic parameters was used to calculate the double profile. To calculate the velocity, energy and trajectory of the kinks, the block method was used. The results obtained made it possible to explain in which region of the plasmid the formation of a transcription bubble is most likely, as well to understand in which direction the bubble will move and the transcription process will go.

| [1] | Yakushevich LV (2006) Nonlinear physics of DNA Weinheim: Wiley. |
| [2] | Dauxois T, Peyrard M (2006) Physics of Solitons Cambridge: Cambridge University Press. |
| [3] | Shapovalov AV, Krasnobaeva LA (2009) Solitons of the sine-Gordon equation Tomsk: TGU, Available from: http://vital.lib.tsu.ru/vital/access/manager/Repository/vtls:000392302. |
| [4] | Ivancevic VG, Ivancevic TT (2013) Sine-Gordon solitons, kinks and breathers as physical models of nonlinear excitations in living cellular structures. J Geom Symmetry Phys 31: 1-56. |
| [5] |
Englander SW, Kallenbach NR, Heeger AJ, et al. (1980) Nature of the open state in long polynucleotide double helices: possibility of soliton excitations. Proc Natl Acad Sci USA 77: 7222-7226. doi: 10.1073/pnas.77.12.7222

|
| [6] |
Forth S, Sheinin MY, Inman J, et al. (2013) Torque measurement at the single-molecule level. Annu Rev Biophys 42: 583-604. doi: 10.1146/annurev-biophys-083012-130412
|
| [7] | Severin ES (2020) Biochemistry Moscow: GEOTAR–Media, Available from: https://expose.gpntbsib.ru/expose/vnp-4f32781c/book/Е2019-3257830172062. |
| [8] |
Salerno M (1995) Nonlinear dynamics of plasmid pBR322 promoters. Nonlinear Excitations in Biomolecules New York: Springer, 147-153. doi: 10.1007/978-3-662-08994-1_10
|
| [9] | Yakushevich LV, Krasnobaeva LA, Shapovalov AV, et al. (2005) One-and two-soliton solutions of the sine-Gordon equation as applied to DNA. Biophysics 50: 450-455. |
| [10] |
Yakushevich LV, Krasnobaeva LA (2016) Forced oscillations of DNA bases. Biophysics 61: 241-250. doi: 10.1134/S000635091602024X
|
| [11] |
Grinevich AA, Ryasik AA, Yakushevich LV (2015) Trajectories of DNA bubbles. Chaos, Soliton Fract 75: 62-75. doi: 10.1016/j.chaos.2015.02.009
|
| [12] |
Yakushevich LV, Krasnobaeva LA (2019) Plasmid pBR322 and nonlinear conformational distortions (kinks). Math Biol Bioinform 14: 327-339. doi: 10.17537/2019.14.327
|
| [13] |
Watson N (1988) A new revision of the sequence of plasmid pBR322. Gene 70: 399-403. doi: 10.1016/0378-1119(88)90212-0
|
| [14] | The pBR322 DNA sequence Available from: http://www.ncbi.nlm.nih.gov/nuccore/J01749.1. |
| [15] |
Krasnobaeva LA, Yakushevich LV (2015) Rotational dynamics of bases in the gene coding interferon alpha 17 (IFNA17). J Bioinf Comput Biol 13: 1540002. doi: 10.1142/S0219720015400028
|
| [16] |
Yakushevich LV, Krasnobaeva LA (2008) A new approach to studies of non-linear dynamics of kinks activated in inhomogeneous polynucleotide chains. Int J Nonlin Mech 43: 1074-1081. doi: 10.1016/j.ijnonlinmec.2008.05.003
|
| [17] |
Yakushevich L (2017) On the mechanical analogue of DNA. J Biol Phys 43: 113-125. doi: 10.1007/s10867-016-9437-0
|
| [18] |
Yakushevich LV, Savin AV, Manevitch LI (2002) Nonlinear dynamics of topological solitons in DNA. Phys Rev E 66: 016614. doi: 10.1103/PhysRevE.66.016614
|
| [19] |
McLaughlin DW, Scott AC (1978) Perturbation analysis of fuxon dynamics. Phys Rev A 18: 1652-1658. doi: 10.1103/PhysRevA.18.1652
|
| [20] |
Yakushevich LV, Krasnobaeva LA (2007) Effects of dissipation and external fields on the dynamics of conformational distortions in DNA. Biophysics 52: 237-243. doi: 10.1134/S0006350907020066
|
| [21] |
Yakushevich LV, Ryasik AA (2012) Dynamical characteristics of DNA kinks and antikinks. Comput Res Model 4: 209-217. doi: 10.20537/2076-7633-2012-4-1-209-217
|
 biophy-08-02-016-s001.pdf biophy-08-02-016-s001.pdf |
 |






Figures(7) / Tables(2)

Ludmila V. Yakushevich, Larisa A. Krasnobaeva. Double energy profile of pBR322 plasmid[J]. AIMS Biophysics, 2021, 8(2): 221-232. doi: 10.3934/biophy.2021016







 DownLoad:
DownLoad: