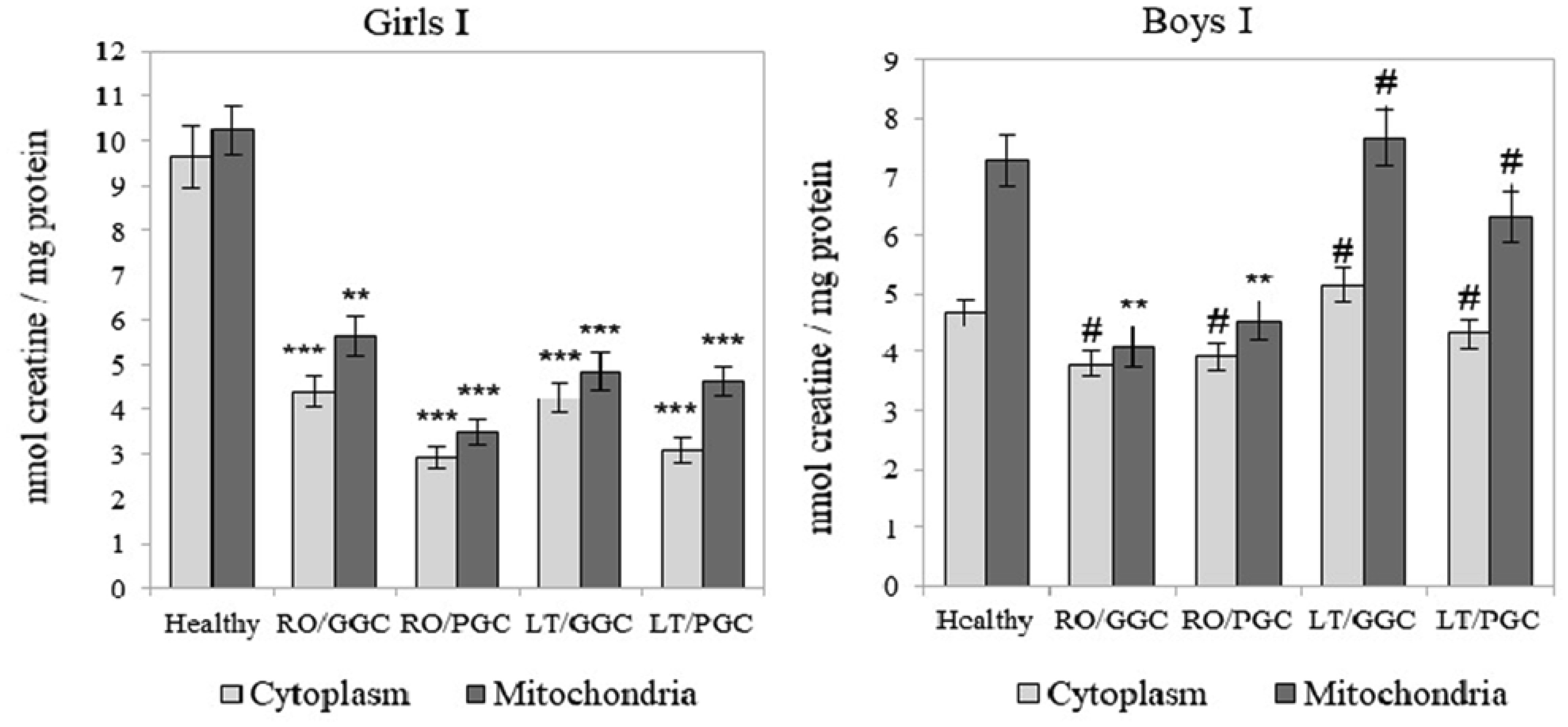
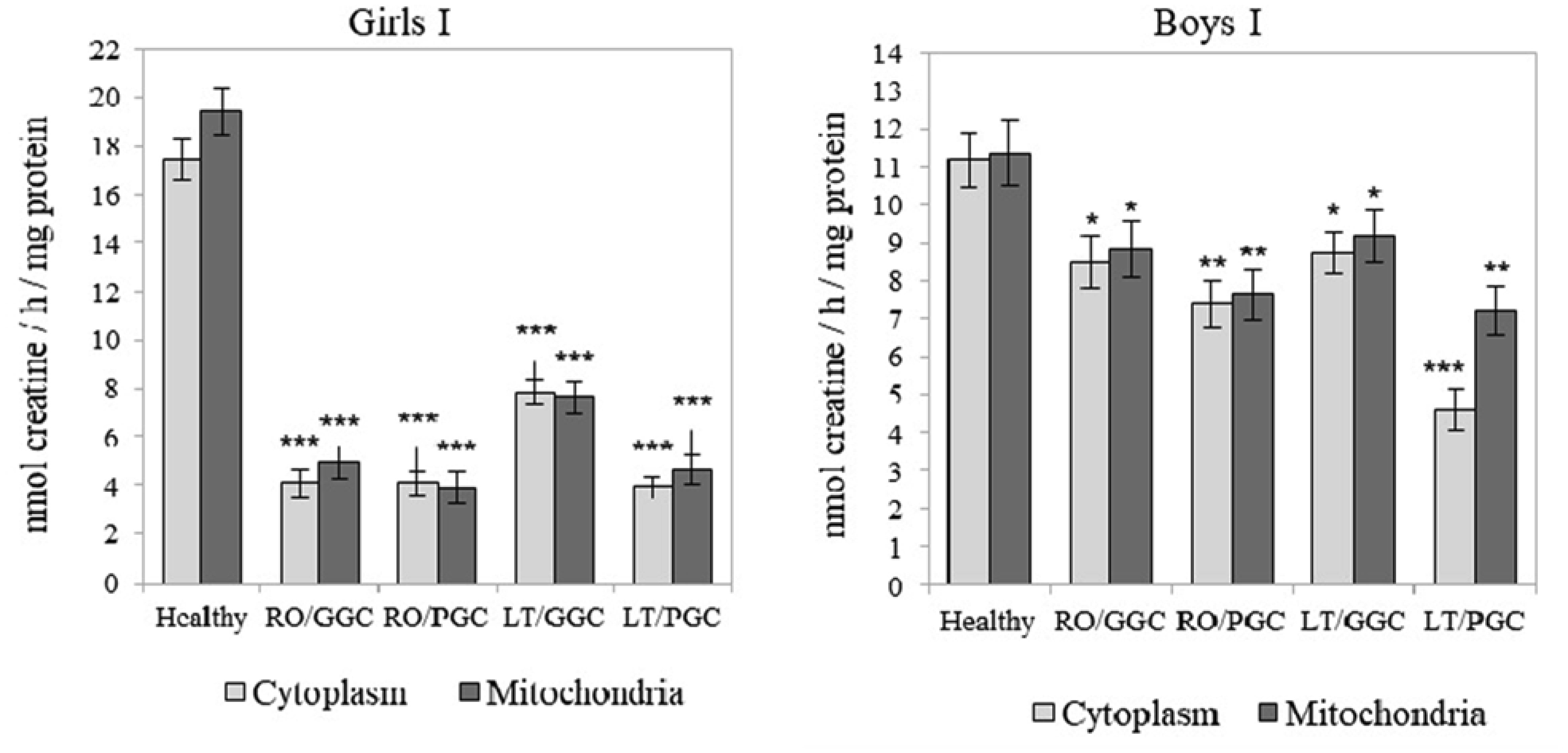
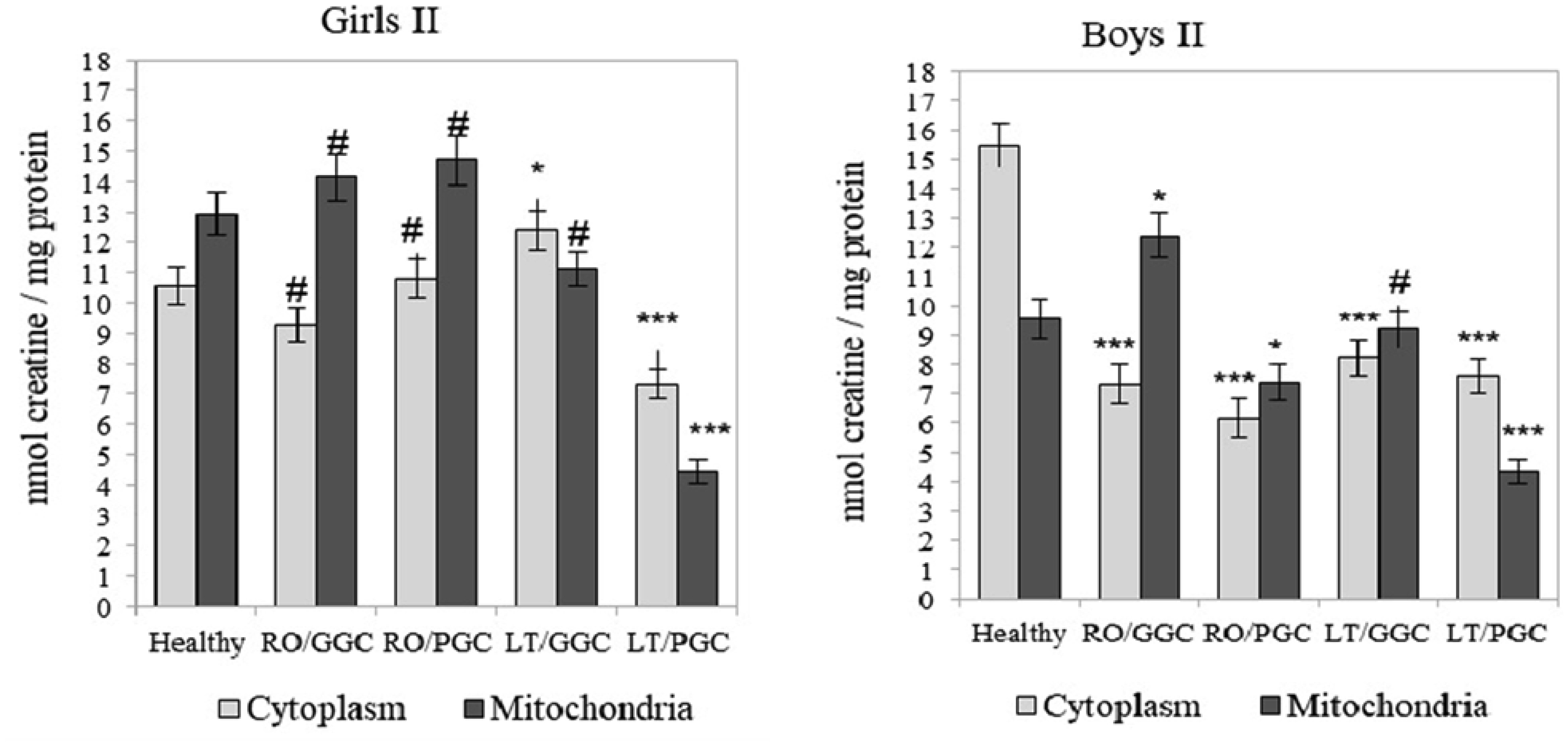
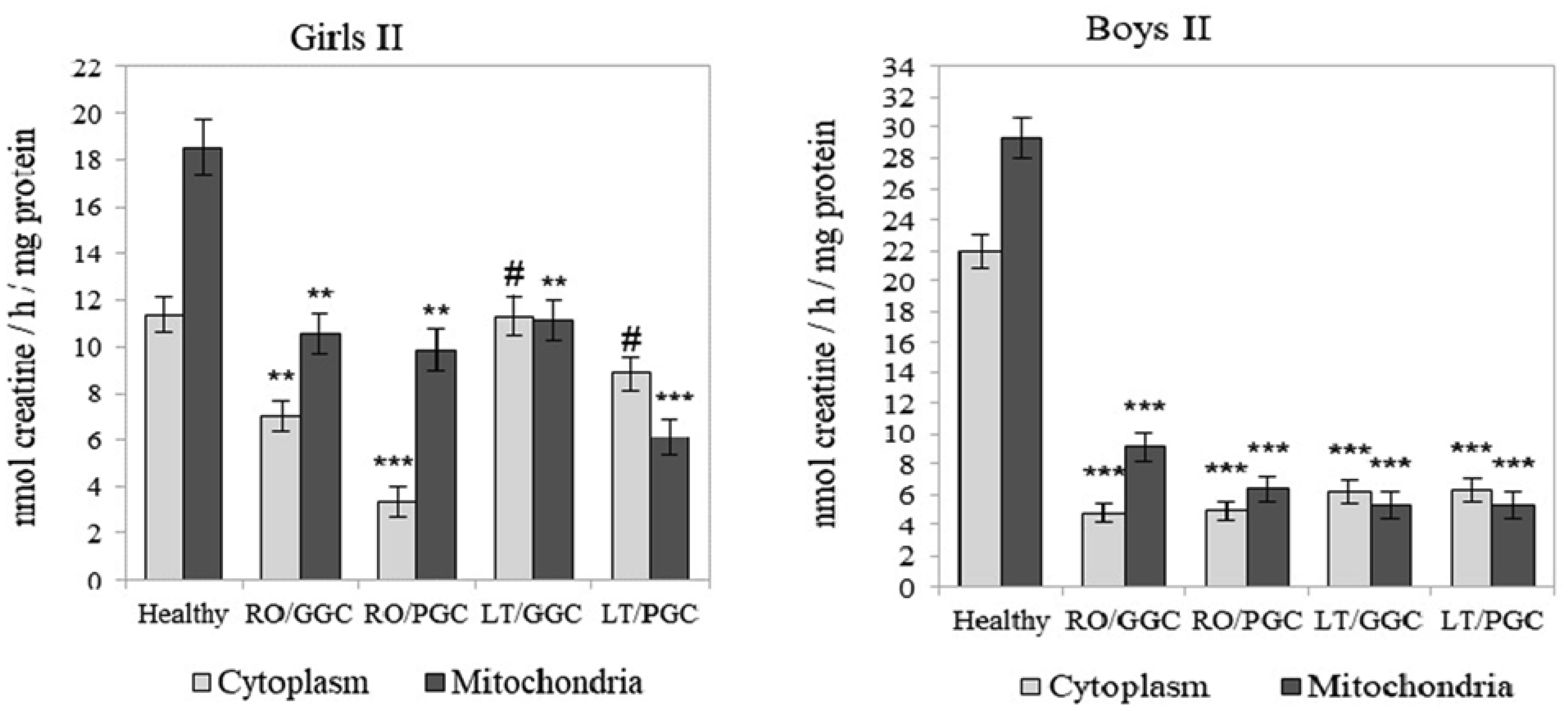
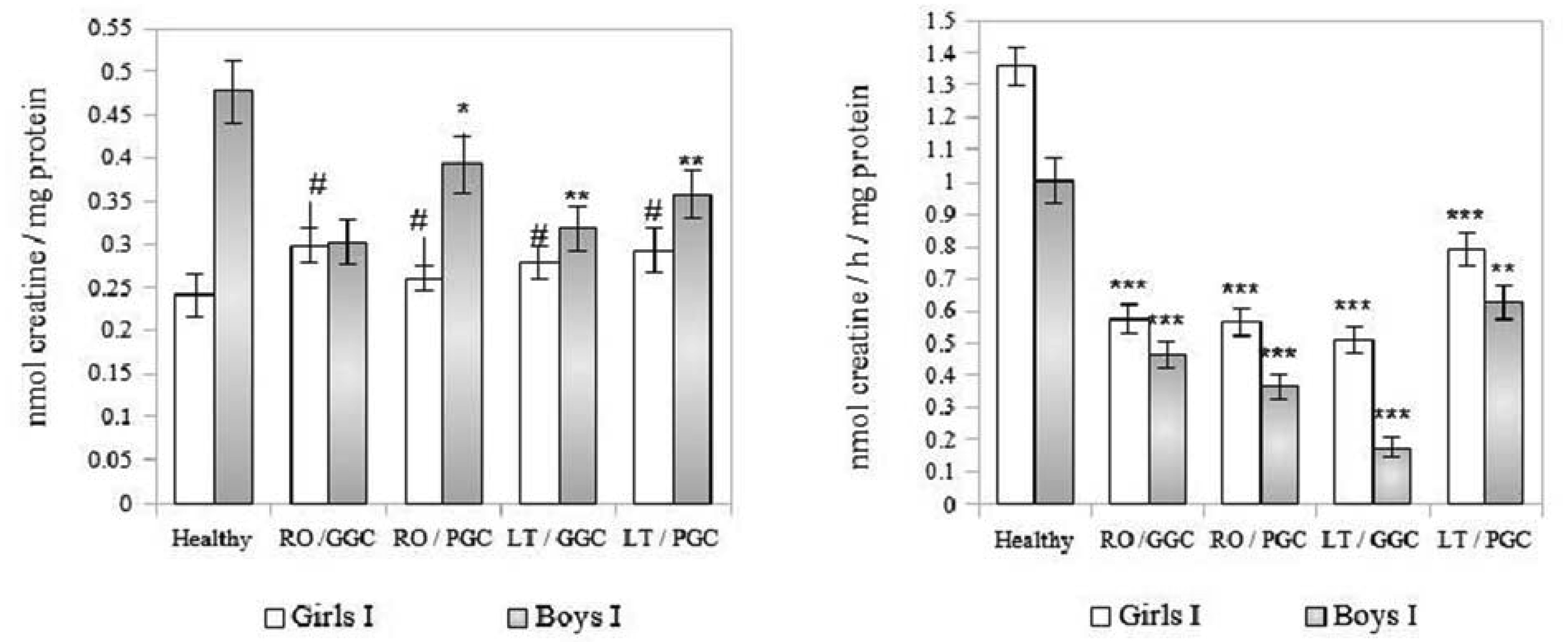
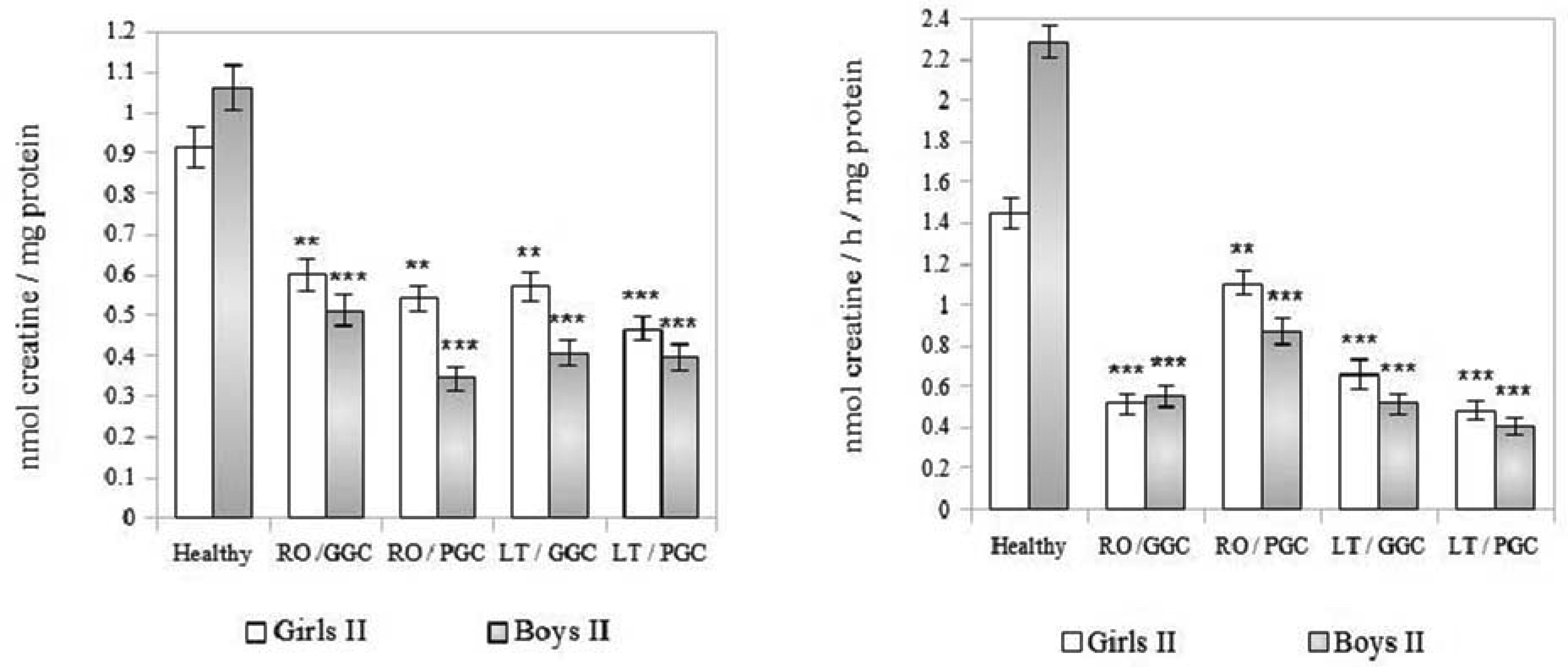
The creatine (Cr)/creatine kinase (CK) system plays a crucial role in cellular energy metabolism, glucose control, and the immune response. This study aimed to investigate this system in the peripheral blood leukocytes (PBL) of Armenians with type 1 diabetes (T1D) who received insulin therapy. A total of 270 Armenian participants were enrolled in the study and were divided into two age groups: Group I - included children and preadolescents; and Group II - included adolescents and young adults. Within each group, the participants were further categorized based on sex and disease duration: Recent-onset T1D (RO-T1D), and Long-term T1D (LT-T1D). A group of healthy individuals that were matched for age and sex served as controls. The glycemic control (GC) was assessed using the glycated hemoglobin (HbA1c) test. In girls of Group I with LT-T1D, both the Cr levels and CK activity were significantly reduced in both the cytoplasm and mitochondria of the PBL, despite having a good GC. In contrast, age-matched boys showed a relatively stable Cr metabolism. Beginning at puberty, only girls with LT-T1D and a poor GC showed decreased Cr levels and a reduced CK activity, which was most pronounced in the mitochondria. In boys of Group II, the cytoplasmic Cr levels decreased by half, regardless of the GC or disease duration, while the mitochondrial Cr levels decreased by 55% only in boys with LT-T1D and a poor GC. Their CK activity decreased by 80% in both cellular compartments, regardless of the GC and duration of diabetes. Notably, changes in the plasma were less specific. Our findings indicate age- and sex-dependent changes in Cr metabolism in the cytoplasm and mitochondria of PBL in Armenians with T1D, which are influenced by the glycemic status and the disease duration. Further extensive studies in this area may provide insights into potential strategies to control metabolism in T1D to counteract autoimmunity and immune dysregulation, and may also serve as a basis for the development of targeted therapeutic interventions.
Citation: Nina Alchujyan, Elena Aghajanova, Nina Movsesyan, Arthur Melkonyan, Artashes Guevorkian, Armen Andreasyan, Margarita Hovhannisyan. Sex-specific alterations in creatine metabolism in cellular compartments of peripheral blood leukocytes in type 1 diabetes[J]. AIMS Bioengineering, 2024, 11(4): 600-616. doi: 10.3934/bioeng.2024028
The creatine (Cr)/creatine kinase (CK) system plays a crucial role in cellular energy metabolism, glucose control, and the immune response. This study aimed to investigate this system in the peripheral blood leukocytes (PBL) of Armenians with type 1 diabetes (T1D) who received insulin therapy. A total of 270 Armenian participants were enrolled in the study and were divided into two age groups: Group I - included children and preadolescents; and Group II - included adolescents and young adults. Within each group, the participants were further categorized based on sex and disease duration: Recent-onset T1D (RO-T1D), and Long-term T1D (LT-T1D). A group of healthy individuals that were matched for age and sex served as controls. The glycemic control (GC) was assessed using the glycated hemoglobin (HbA1c) test. In girls of Group I with LT-T1D, both the Cr levels and CK activity were significantly reduced in both the cytoplasm and mitochondria of the PBL, despite having a good GC. In contrast, age-matched boys showed a relatively stable Cr metabolism. Beginning at puberty, only girls with LT-T1D and a poor GC showed decreased Cr levels and a reduced CK activity, which was most pronounced in the mitochondria. In boys of Group II, the cytoplasmic Cr levels decreased by half, regardless of the GC or disease duration, while the mitochondrial Cr levels decreased by 55% only in boys with LT-T1D and a poor GC. Their CK activity decreased by 80% in both cellular compartments, regardless of the GC and duration of diabetes. Notably, changes in the plasma were less specific. Our findings indicate age- and sex-dependent changes in Cr metabolism in the cytoplasm and mitochondria of PBL in Armenians with T1D, which are influenced by the glycemic status and the disease duration. Further extensive studies in this area may provide insights into potential strategies to control metabolism in T1D to counteract autoimmunity and immune dysregulation, and may also serve as a basis for the development of targeted therapeutic interventions.

| [1] | Atkinson MA, Eisenbarth GS, Michels AW (2014) Type 1 diabetes. Lancet 383: 69-82. https://doi.org/10.1016/S0140-6736(13)60591-7 |
| [2] | Mobasseri M, Shirmohammadi M, Amiri T, et al. (2020) Prevalence and incidence of type 1 diabetes in the world: a systematic review and meta-analysis. Health Promot Perspe 10: 98-115. https://doi.org/10.34172/hpp.2020.18 |
| [3] | Navasardyan LV (2014) Epidemiology of type 1 diabetes in children and adolescents in the Republic of Armenia. New Arm Med 8: 89-94. Available from: https://ysmu.am/website/documentation/files/79650ab6.pdf |
| [4] | Troullinaki M, Chen LS, Witt A, et al. (2020) Robo4-mediated pancreatic endothelial integrity decreases inflammation and islet destruction in autoimmune diabetes. FASEB J 34: 3336-3346. https://doi.org/10.1096/fj.201900125RR |
| [5] | Babon JA, DeNicola ME, Blodgett DM, et al. (2016) Analysis of self-antigen specificity of islet-infiltrating T cells from human donors with type 1 diabetes. Nat Med 22: 1482-1487. https://doi.org/10.1038/nm.4203 |
| [6] | Schlattner U, Klaus A, Ramirez Rios S, et al. (2016) Cellular compartmentation of energy metabolism: creatine kinase microcompartments and recruitment of B-type creatine kinase to specific subcellular sites. Amino Acids 48: 175-1774. https://doi.org/10.1007/s00726-016-2267-3 |
| [7] | Solis MY, Artioli GG, Gualano B (2021) Potential of creatine in glucose management and diabetes. Nutrients 13: 570-583. https://doi.org/10.3390/nu13020570 |
| [8] | Bredahl EC, Eckerson JM, Tracy SM, et al. (2021) The role of creatine in the development and activation of immune responses. Nutrients 13: 751-768. https://doi.org/10.3390/nu13030751 |
| [9] | World Medical Association, Declaration of Helsinki, Ethical Principles for Medical Research Involving Human Subjects. 59th WMA General Assembly, Seoul, October 2008 . |
| [10] | Cengiz E, Danne T, Ahmad T, et al. (2022) Clinical Practice Consensus Guidelines 2022: Insulin treatment in children and adolescents with diabetes. ISPAD Pediatr Diabetes 23: 1277-1296. https://doi.org/10.1111/pedi.13442 |
| [11] | Chang J, Hoke C, Ettinger B, et al. (1998) Evaluation and interference study of hemoglobinA1c measured by turbidimetric inhibition immunoassay. Am J Clin Pathol 109: 274-278. https://doi.org/10.1093/ajcp/109.3.274 |
| [12] | Ivanov II, Korovkin BF, Markelov IM (1974) Introduction to clinical enzymology. Leningrad: Medicine 134-136. |
| [13] | Lowry OH, Rosebrough NJ, Farr AL, et al. (1951) Protein measurement with the Folin phenol reagent. J Biol Chem 193: 265-275. https://doi.org/10.1016/S0021-9258(19)52451-6 |
| [14] | Wei N, Zheng H, Nathan DM (2014) Empirically establishing blood glucose targets to achieve HbA1c goals. Diabetes Care 37: 1048-1051. https://doi.org/10.2337/dc13-2173 |
| [15] | Wallimann T, Tokarska-Schlattner M, Schlattner U (2011) The creatine kinase system and pleiotropic effects of creatine. Amino Acids 40: 1271-1296. https://doi.org/10.1007/s00726-011-0877-3 |
| [16] | Kazak L, Cohen P (2020) Creatine metabolism: energy homeostasis, immunity and cancer biology. Nat Rev Endocrinol 16: 421-436. https://doi.org/10.1038/s41574-020-0365-5 |
| [17] | Brewster LM (2018) Creatine kinase, energy reserve, and hypertension: from bench to bedside. Ann Transl Med 6: 292. https://doi.org/10.21037/atm.2018.07.15 |
| [18] | de la Cuesta-Zuluaga J, Kelley ST, Chen Y, et al. (2019) Age-and sex-dependent patterns of gut microbial diversity in human adults. Msystems 4: 1-12. https://doi.org/10.1128/mSystems.00261-19 |
| [19] | Klein SL, Flanagan KL (2016) Sex differences in immune responses. Nat Rev Immunol 16: 626-638. https://doi.org/10.1038/nri.2016.90 |
| [20] | Alves CR, Ferreira JC, de Siqueira-Filho MA, et al. (2012) Creatine-induced glucose uptake in type 2 diabetes: a role for AMPK-alpha?. Amino Acids 43: 1803-1807. https://doi.org/10.1007/s00726-012-1246-6 |
| [21] | Taii A, Tachikawa M, Ohta Y, et al. (2020) Determination of intrinsic creatine transporter (Slc6a8) activity and creatine transport function of leukocytes in rats. Biol Pharm Bull 43: 474-479. https://doi.org/10.1248/bpb.b19-00807 |
| [22] | Frederiksen H, Johannsen TH, Andersen SE, et al. (2020) Sex-specific estrogen levels and reference intervals from iInfancy to late adulthood determined by LC-MS/MS. J Clin Endocrinol Metab 105: 754-768. https://doi.org/10.1210/clinem/d+gz196 |
| [23] | Ji L, Zhao X, Zhang B, et al. (2019) Slc6a8-mediated creatine uptake and accumulation reprogram macrophage polarization via regulating cytokine responses. Immunity 51: 272-284. https://doi.org/10.1016/j.immuni.2019.06.007 |
| [24] | Samborska B, Roy DG, Rahbani JF, et al. (2022) Creatine transport and creatine kinase activity is required for CD8+T cell immunity. Cell Rep 38: 110446. https://doi.org/10.1016/j.celrep.2022.110446 |
| [25] | Post A, Groothof D, Schutten JC, et al. (2021) Plasma creatine and incident type 2 diabetes in a general population-based cohort: the prevend study. Clin Endocrinol 94: 563-567. https://doi.org/10.1111/cen.14396 |
| [26] | Dolder M, Walzel B, Speer O, et al. (2003) Inhibition of the mitochondrial permeability transition by creatine kinase substrates. Requirement for microcompartmentation. J Biol Chem 278: 17760-17766. https://doi.org/10.1074/jbc.M208705200 |
| [27] | Safdar A, Yardley NJ, Snow R, et al. (2008) Global and targeted gene expression and protein content in skeletal muscle of young men following short-term creatine monohydrate supplementation. Physiol Genom 32: 219-228. https://doi.org/10.1152/physiolgenomics.00157.2007 |
| [28] | Deldicque L, Louis M, Theisen D, et al. (2005) Increased IGF mRNA in human skeletal muscle after creatine supplementation. Med Sci Sports Exerc 37: 731-736. https://doi.org/10.1249/01.MSS.0000162690.39830.27 |
| [29] | Šimonienė D, Platukiene A, Prakapienė E, et al. (2020) Insulin resistance in type 1 diabetes mellitus and its association with patient's micro- and macrovascular complications, sex hormones, and other clinical data. Diabetes Ther 11: 161-174. https://doi.org/10.1007/s13300-019-00729-5 |
| [30] | Sharma G, Prossnitz ER (2017) G-protein-coupled estrogen receptor and sex-specific metabolic homeostasis. Adv Exp Med Biol 1043: 427-453. https://doi.org/10.1007/978-3-319-70178-3_20 |
| [31] | Tao Z, Cheng Z (2023) Hormonal regulation of metabolism-recent lessons learned from insulin and estrogen. Clin Sci 137: 415-434. https://doi.org/10.1042/CS20210519 |
| [32] | Khan D, Ansar Ahmed S (2016) The immune system is a natural target for estrogen action: opposing effects of estrogen in two prototypical autoimmune diseases. Front Immunol 6: 635-643. https://doi.org/10.3389/fimmu.2015.00635 |
| [33] | Kostopoulou E, Livada I, Partsalaki I, et al. (2020) The role of carbohydrate counting in glycemic control and oxidative stress in patients with type 1 diabetes mellitus (T1DM). Hormones 19: 433-438. https://doi.org/10.1007/s42000-020-00189-8 |
| [34] | Brosnan ME, Brosnan JT (2016) The role of dietary creatine. Amino Acids 48: 1785-1791. https://doi.org/10.1007/s00726-016-2188-1 |
| [35] | González P, Lozano P, Ros G, et al. (2023) Hyperglycemia and oxidative stress: an integral, updated and critical overview of their metabolic interconnections. Int J Mol Sci 24: 9352. https://doi.org/10.3390/ijms24119352 |
| [36] | Caldwell RW, Rodriguez PC, Toque HA, et al. (2018) Arginase: a multifaceted enzyme important in health and disease. Physiol Rev 98: 641-665. https://doi.org/10.1152/physrev.00037.2016 |
| [37] | Genet S, Kale RK, Baquer NZ (2000) Effects of free radicals on cytosolic creatine kinase activities and protection by antioxidant enzymes and sulfhydryl compounds. Mol Cell Biochem 210: 23-28. https://doi.org/10.1023/a:1007071617480 |
| [38] | Yuan G, Kaneko M, Masuda H, et al. (1992) Decrease in heart mitochondrial creatine kinase activity due to oxygen free radicals. Biochim Biophys Acta 1140: 78-84. https://doi.org/10.1016/0005-2728(92)90022-t |
| [39] | Ren Y, Li Z, Li W, et al. (2022) Arginase: biological and therapeutic implications in dabetes mellitus and its complications. Oxid Med Cell Longev 2022: 2419412. https://doi.org/10.1155/2022/2419412 |
| [40] | Alchujyan NKh, Hovhannisyan MR, Movsesyan NH, et al. (2021) Sexual dimorphism in alternative metabolic pathways of L-arginine in circulating leukocytes in young people with type 1 diabetes mellitus. Endocrine Res 46: 149-159. https://doi.org/10.1080/07435800.2021.1920608 |
| [41] | Tomita Y, Cagnone G, Fu Z, et al. (2021) Vitreous metabolomics profiling of proliferative diabetic retinopathy. Diabetologia 64: 70-82. https://doi.org/10.1007/s00125-020-05309-y |
| [42] | Ingoglia F, Chong JL, Pasquali M, et al. (2021) Creatine metabolism in patients with urea cycle disorders. Mol Genet Metab Rep 29: 100791. https://doi.org/10.1016/j.ymgmr.2021.100791 |
| [43] | Kietadisorn R, Juni RP, Moens AL (2012) Tackling endothelial dysfunction by modulating NOS uncoupling: new insights into its pathogenesis and therapeutic possibilities. Am J Physiol Endocrinol Metab 302: E481-E495. https://doi.org/10.1152/ajpendo.00540.2011 |
| [44] | Ahmad R, Hussain A, Ahsan H (2019) Peroxynitrite: cellular pathology and implications in autoimmunity. J Immunoassay Immunochem 40: 123-138. https://doi.org/10.1080/15321819.2019.1583109 |
| [45] | Delwing D, Cornélio AR, Wajner M, et al. (2007) Arginine administration reduces creatine kinase activity in rat cerebellum. Metab Brain Dis 22: 13-23. https://doi.org/10.1007/s11011-006-9028-z |
| [46] | Darabedian N, Ji W, Fan M, et al. (2023) Depletion of creatine phosphagen energetics with a covalent creatine kinase inhibitor. Nat Chem Biol 19: 815-824. https://doi.org/10.1038/s41589-023-01273-x |
| [47] | Wang W, Lee J, Hao H, et al. (2017) Hydrogen peroxide (H2O2) irreversibly inactivates creatine kinase from Pelodiscus sinensis by targeting the active site cysteine. Int J Biol Macromol 105: 1595-1601. https://doi.org/10.1016/j.ijbiomac.2017.03.025 |
| [48] | Cheprasova AA, Popov SS, Pashkov AN, et al. (2022) Parameters of oxidative stress and activity of antioxidant enzymes in the saliva of patients with type 1 diabetes mellitus. Bull Exp Biol Med 172: 552-557. https://doi.org/10.1007/s10517-022-05431-4 |
| [49] | Gonçalves MG, Medeiros MA, de Lemos LIC, et al. (2022) Effects of creatine supplementation on histological and biochemical parameters in the kidney and pancreas of 7streptozotocin-induced diabetic rats. Nutrients 14: 431-454. https://doi.org/10.3390/nu14030431 |
| [50] | Riesberg LA, McDonald TL, Wang Y, et al. (2018) Creatinine downregulates TNF-α in macrophage and T cell lines. Cytokine 110: 29-38. https://doi.org/10.1016/j.cyto.2018.04.021 |
| [51] | Sironi C, Bodega F, Zocchi L, et al. (2019) Effects of creatine treatment on jejunial phenotypes in a rat model of acidosis. Antioxidants 8: 22. https://doi.org/10.3390/antiox8070225 |
| [52] | Antonio J, Candow DG, Forbes SC, et al. (2021) Common questions and misconceptions about creatine supplementation: what does the scientific evidence really show?. J Int Soc Sports Nutr 18: 13. https://doi.org/10.1186/s12970-021-00412-w |
| [53] | Ostojic SM (2022) Low tissue creatine: a therapeutic target in clinical nutrition. Nutrients 14: 1230-1238. https://doi.org/10.3390/nu14061230 |
| [54] | Takeshita S, Saito Y, Oyama Y, et al. (2021) Infection-associated decrease of serum creatine kinase levels in Fukuyama congenital muscular dystrophy. Brain Dev 43: 440-447. https://doi.org/10.1016/j.braindev.2020.11.009 |
| [55] | McMillan HJ, Lochmüller H (2021) Biomarkers in duchenne and becker muscular dystrophies. Muscle Nerve 64: 4-5. https://doi.org/10.1002/mus.27342 |
Figures(6) / Tables(2)

Nina Alchujyan, Elena Aghajanova, Nina Movsesyan, Arthur Melkonyan, Artashes Guevorkian, Armen Andreasyan, Margarita Hovhannisyan. Sex-specific alterations in creatine metabolism in cellular compartments of peripheral blood leukocytes in type 1 diabetes[J]. AIMS Bioengineering, 2024, 11(4): 600-616. doi: 10.3934/bioeng.2024028







 DownLoad:
DownLoad: