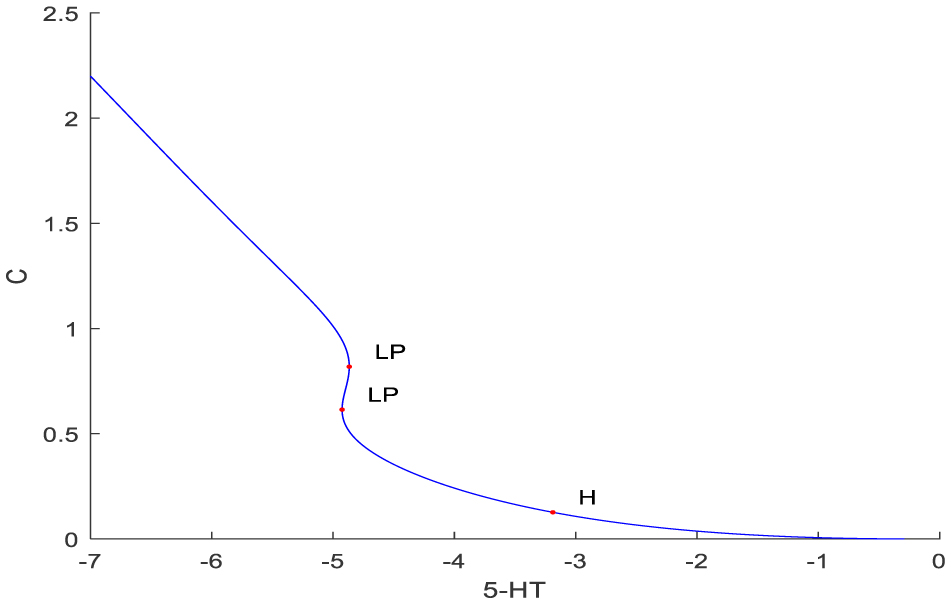
Molecular biological networks are highly nonlinear systems that exhibit limit point singularities. Bifurcation analysis and multiobjective nonlinear model predictive control (MNLMPC) of a molecular network problem represented by the Pettigrew model were performed. The Matlab program MATCONT (Matlab continuation) was used for the bifurcation analysis and the optimization language PYOMO (python optimization modeling objects) was used for performing the multiobjective nonlinear model predictive control. MATCONT identified the limit points, branch points, and Hopf bifurcation points using appropriate test functions. The multiobjective nonlinear model predictive control was performed by first performing single objective optimal control calculations and then minimizing the distance from the Utopia point, which was the coordinate of minimized values of each objective function. The presence of limit points (albeit in an infeasible region) enabled the MNLMPC calculations to result in the Utopia solution. MNLMPC of the partial models also resulted in Utopia solutions.
Citation: Lakshmi N Sridhar. Integration of bifurcation analysis and optimal control of a molecular network[J]. AIMS Bioengineering, 2024, 11(2): 266-280. doi: 10.3934/bioeng.2024014
Molecular biological networks are highly nonlinear systems that exhibit limit point singularities. Bifurcation analysis and multiobjective nonlinear model predictive control (MNLMPC) of a molecular network problem represented by the Pettigrew model were performed. The Matlab program MATCONT (Matlab continuation) was used for the bifurcation analysis and the optimization language PYOMO (python optimization modeling objects) was used for performing the multiobjective nonlinear model predictive control. MATCONT identified the limit points, branch points, and Hopf bifurcation points using appropriate test functions. The multiobjective nonlinear model predictive control was performed by first performing single objective optimal control calculations and then minimizing the distance from the Utopia point, which was the coordinate of minimized values of each objective function. The presence of limit points (albeit in an infeasible region) enabled the MNLMPC calculations to result in the Utopia solution. MNLMPC of the partial models also resulted in Utopia solutions.

| [1] | Smolen P, Baxter DA, Byrne JH (2000) Mathematical modelling of gene networks. Neuron 26: 567-580. https://doi.org/10.1016/S0896-6273(00)81194-0 |
| [2] | Goldsmith JR, Byrne JH (1993) Bag cell extract inhibits tail-siphon withdrawal reflex, suppresses long-term but not short-term sensitization, and attenuates sensory-to-motor neuron synapses in Aplysia. J Neurosci 13: 1688-1700. https://doi.org/10.1523/JNEUROSCI.13-04-01688.1993 |
| [3] | Goldbeter A (2002) Computational approaches to cellular rhythms. Nature 420: 238-245. https://doi.org/10.1038/nature01259 |
| [4] | Kitano H (2002) Systems biology: A brief overview. Science 295: 1662-1664. https://doi.org/10.1126/science.1069492 |
| [5] | Song H, Smolen P, Av-Ron E, et al. (2006) Bifurcation and singularity analysis of a molecular network for the induction of long-term memory. Biophys J 90: 2309-2325. https://doi.org/10.1529/biophysj.105.074500 |
| [6] | Sridhar LN (2023) Multi objective nonlinear model predictive control of diabetes models considering the effects of insulin and exercise. Archives Clin Med Microbiol 2: 60-69. https://doi.org/10.33140/ACMMJ |
| [7] | Pinsker H, Carew TJ, Hening W, et al. (1973) Long-term sensitization of a defensive withdrawal reflex in Aplysia californica. Science 182: 1039-1042. https://doi.org/10.1126/science.182.4116.1039 |
| [8] | Frost WN, Castelluci VF, Hawkins RD, et al. (1985) Monosynaptic connections made by the sensory neurons of the gill and siphon-withdrawal reflex in Aplysia participate in the storage of long-term memory for sensitization. Proc Natl Acad Sci 82: 8266-8269. https://doi.org/10.1073/pnas.82.23.8266 |
| [9] | Walters ET (1987) Site-specific sensitization of defensive reflexes in Aplysia: A simple model of long-term hyperalgesia. J Neurosci 7: 400-407. https://doi.org/10.1523/jneurosci.07-02-00400.1987 |
| [10] | Walters ET (1987) Multiple sensory neuronal correlates of site-specific sensitization in Aplysia. J Neurosci 7: 408-417. https://doi.org/10.1523/jneurosci.07-02-00400.1987 |
| [11] | Castellucci VF, Blumenfeld H, Goelet P, et al. (1989) Inhibitor of protein synthesis blocks long-term behavioral sensitization in the isolated gill-withdrawal reflex of Aplysia. J Neurobiol 20: 1-9. https://doi.org/10.1002/neu.480200102 |
| [12] | Cleary LJ, Lee WL, Byrne JH (1998) Cellular correlates of long-term sensitization in Aplysia. J Neurosci 18: 5988-5998. https://doi.org/10.1101/lm.045450.117 |
| [13] | Wright WG, McCance EF, Carew TJ (1996) Developmental emergence of long-term memory for sensitization in Aplysia. Neurobiol Learn Mem 65: 261-268. https://doi.org/10.1006/Nlme.1996.0031 |
| [14] | Levenson J, Byrne JH, Eskin A (1999) Levels of serotonin in the hemolymph of Aplysia are modulated by light/dark cycles and sensitization training. J Neurosci 19: 8094-8103. https://doi.org/10.1523/JNEUROSCI.19-18-08094.1999 |
| [15] | Sutton MA, Ide J, Masters SE, et al. (2002) Interaction between amount and pattern of training in the induction of intermediate- and long-term memory for sensitization in Aplysia. Learn Mem 9: 29-40. https://doi.org/10.1101/lm.44802 |
| [16] | Wainwright ML, Byrne JH, Cleary LJ (2004) Dissociation of morphological and physiological changes associated with long-term memory in Aplysia. J Neurophysiol 92: 2628-2632. https://doi.org/10.1152/jn.00335.2004 |
| [17] | Wainwright ML, Zhang H, Byrne JH, et al. (2002) Localized neuronal outgrowth induced by long-term sensitization training in Aplysia. J Neurosci 22: 4132-4141. https://doi.org/10.1523/JNEUROSCI.22-10-04132.2002 |
| [18] | Pettigrew D, Smolen P, Baxter DA, et al. (2005) Dynamic properties of regulatory motifs associated with induction of three temporal domains of memory in Aplysia. J Comput Neurosci 18: 163-181. https://doi.org/10.1007/s10827-005-6557-0 |
| [19] | Song H, Smolen P, Av-Ron E, et al. (2006) Bifurcation and singularity analysis of a molecular network for the induction of long-term memory. Biophys J 90: 2309-2325. https://doi.org/10.1529/biophysj.105.074500 |
| [20] | Dhooge A, Govearts W, Kuznetsov AY (2003) MATCONT: A MATLAB package for numerical bifurcation analysis of ODEs. ACM T Math Software 29: 141-164. https://doi.org/10.1145/779359.779362 |
| [21] | Dhooge A, Govaerts W, Kuznetsov YA, et al. (2003) CL_MATCONT: A continuation toolbox in Matlab. Proceedings of the 2003 ACM Symposium on Applied Computing : 161-166. https://doi.org/10.1145/952532.952567 |
| [22] | Kuznetsov YA (1998) Elements of Applied Bifurcation Theory. New York: Springer. |
| [23] | Kuznetsov YA (2009) Five Lectures on Numerical Bifurcation Analysis. Utrecht University NL. |
| [24] | Govaerts WJF (2000) Numerical Methods for Bifurcations of Dynamical Equilibria. Society for Industrial and Applied Mathematics. |
| [25] | Flores-Tlacuahuac A, Morales P, Riveral-Toledo M Multiobjective nonlinear model predictive control of a class of chemical reactors. Ind Eng Chem Res 51: 5891-5899. https://doi.org/10.1021/ie201742e |
| [26] | Sridhar LN (2021) Single and multiobjective optimal control of epidemic models involving vaccination and treatment. J Biostat Epidemiol 7: 25-35. https://doi.org/10.18502/jbe.v7i1.6292 |
| [27] | Sridhar LN (2023) Bifurcation analysis and optimal control of the Crowley Martin phytoplankton-zooplankton model that considers the impact of nanoparticles. Explo Mater Sci Res 5: 54-60. https://dx.doi.org/10.47204/EMSR.5.1.2023.054-060 |
| [28] | Sridhar LN (2020) Multiobjective nonlinear model predictive control of pharmaceutical batch crystallizers. Drug Dev Ind Pharm 46: 2089-2097. https://doi.org/10.1080/03639045.2020.1847135 |
| [29] | Sridhar LN (2019) Multiobjective optimization and nonlinear model predictive control of the continuous fermentation process involving Saccharomyces Cerevisiae. Biofuels 13: 249-264. https://doi.org/10.1080/17597269.2019.1674000 |
| [30] | Sridhar LN (2022) Single and multiobjective optimal control of the wastewater treatment process. Trans Indian Natl Acad Eng 7: 1339-1346. https://doi.org/10.1007/s41403-022-00368-6 |
| [31] | Younis A, Dong Z (2023) Adaptive surrogate assisted multi-objective optimization approach for highly nonlinear and complex engineering design problems. Appl Soft Comput 150: 111065. https://doi.org/10.1016/j.asoc.2023.111065 |
| [32] | Younis A, Dong Z (2022) High-fidelity surrogate based multi-objective optimization algorithm. Algorithms 15: 279. https://doi.org/10.3390/a15080279 |
| [33] | Younis A, Dong Z (2010) Trends, features, and tests of common and recently introduced global optimization methods. Eng Optimiz 42: 691-718. https://doi.org/10.1080/03052150903386674 |
| [34] | Safari A, Younis A, Wang G, et al. (2015) Development of a metamodel-assisted sampling approach to aerodynamic shape optimization problems. J Mech Sci Technol 29: 2013-2024. https://doi.org/10.1007/s12206-015-0422-5 |
| [35] | Miettinen K (1999) Nonlinear Multiobjective Optimization. Berlin: Springer Science & Business Media. |
| [36] | Hart WE, Laird CD, Watson JP, et al. (2017) Pyomo–Optimization Modeling in Python. Berlin: Springer. |
| [37] | Biegler LT (2007) An overview of simultaneous strategies for dynamic optimization. Chem Eng Process 46: 1043-1053. https://doi.org/10.1016/j.cep.2006.06.021 |
| [38] | Wächter A, Biegler L (2006) On the implementation of an interior-point filter line-search algorithm for large-scale nonlinear programming. Math Program 106: 25-57. https://doi.org/10.1007/s10107-004-0559-y |
| [39] | Tawarmalani M, Sahinidis NV (2005) A polyhedral branch-and-cut approach to global optimization. Math Program 103: 225-249. https://doi.org/10.1007/s10107-005-0581-8 |





Figures(6)

Lakshmi N Sridhar. Integration of bifurcation analysis and optimal control of a molecular network[J]. AIMS Bioengineering, 2024, 11(2): 266-280. doi: 10.3934/bioeng.2024014







 DownLoad:
DownLoad: