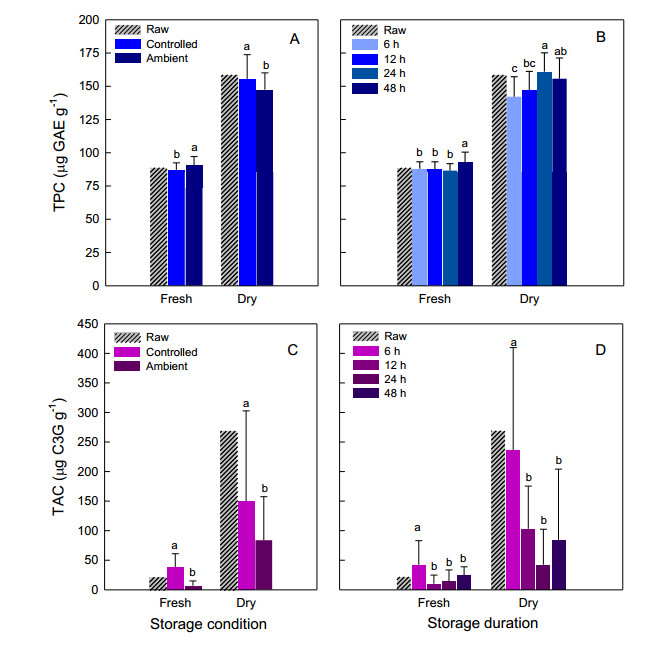
Corn tassel is an agricultural waste product that contains valuable phytochemicals and antioxidants with various potential uses. Proper post-harvest management is vital to maintain the bioactive compounds and favorable properties for processing. This study aimed to evaluate the responses of phenolics, anthocyanins, and antioxidant capacity of purple waxy corn tassels to different storage conditions and durations. Storage conditions (controlled vs. ambient) that varied in temperature and duration (ranging from 6 to 48 hours) significantly altered most of the observed parameters. Phenolics were more resistant to increased temperature and prolonged storage than anthocyanins. Determining the optimal storage duration was slightly complicated as the ideal duration for each observed parameter varied. The tassels can be stored at cold temperatures for up to 48, 6, and 24 hours to obtain the highest levels of phenolics, anthocyanins, and antioxidant activity, respectively. The correlation coefficients between phenolics and antioxidant activity were significant in both fresh and dried tassels. Optimizing the storage conditions to retain phenolics can also help maintain high levels of antioxidant capacity in corn tassels. Controlled storage conditions were the best way to retain tassel weight, phenolics, anthocyanins, and antioxidant capacity in the purple tassels of waxy corn. The most prolonged acceptable storage durations varied depending on the traits. The optimum light and oxygen exposures during storage and the best drying methods are still uncertain; therefore, further research is necessary to establish good handling practices for corn tassels.
Citation: Prakasit Duangpapeng, Abil Dermail, Khundej Suriharn. Phenolics, anthocyanins, and antioxidant capacity in the tassels of purple waxy corn: Effects of temperature and time during storage[J]. AIMS Agriculture and Food, 2024, 9(1): 69-83. doi: 10.3934/agrfood.2024005
Corn tassel is an agricultural waste product that contains valuable phytochemicals and antioxidants with various potential uses. Proper post-harvest management is vital to maintain the bioactive compounds and favorable properties for processing. This study aimed to evaluate the responses of phenolics, anthocyanins, and antioxidant capacity of purple waxy corn tassels to different storage conditions and durations. Storage conditions (controlled vs. ambient) that varied in temperature and duration (ranging from 6 to 48 hours) significantly altered most of the observed parameters. Phenolics were more resistant to increased temperature and prolonged storage than anthocyanins. Determining the optimal storage duration was slightly complicated as the ideal duration for each observed parameter varied. The tassels can be stored at cold temperatures for up to 48, 6, and 24 hours to obtain the highest levels of phenolics, anthocyanins, and antioxidant activity, respectively. The correlation coefficients between phenolics and antioxidant activity were significant in both fresh and dried tassels. Optimizing the storage conditions to retain phenolics can also help maintain high levels of antioxidant capacity in corn tassels. Controlled storage conditions were the best way to retain tassel weight, phenolics, anthocyanins, and antioxidant capacity in the purple tassels of waxy corn. The most prolonged acceptable storage durations varied depending on the traits. The optimum light and oxygen exposures during storage and the best drying methods are still uncertain; therefore, further research is necessary to establish good handling practices for corn tassels.

| [1] | Congress, 117th Session (2021) H.R.4444—Zero Food Waste Act. Available from: https://www.congress.gov/bill/117th-congress/house-bill/4444/text |
| [2] |
Boateng ID (2023) Mechanisms, capabilities, limitations, and economic stability outlook for extracting phenolics from agro-byproducts using emerging thermal extraction technologies and their combinative effects. Food Bioproc Tech 2023: 1–32. https://doi.org/10.1007/s11947-023-03171-5 doi: 10.1007/s11947-023-03171-5

|
| [3] |
Kim HY, Lee KY, Kim M, et al. (2023) A review of the biological properties of purple corn (Zea mays L.). Sci Pharm 91: 6. https://doi.org/10.3390/scipharm91010006 doi: 10.3390/scipharm91010006
|
| [4] |
Ghețe AB, Haș V, Vidican R, et al. (2020) Influence of detasseling methods on seed yield of some parent inbred lines of Turda maize hybrids. Agronomy 10: 729. https://doi.org/10.3390/agronomy10050729 doi: 10.3390/agronomy10050729
|
| [5] | MacRobert JF, Setimela P, Gethi J, et al. (2014) Maize Hybrid Seed Production Manual. CIMMYT, Mexico. |
| [6] |
Mohsen SM, Ammar ASM (2009) Total phenolic contents and antioxidant activity of corn tassel extracts. Food Chem 112: 595–598. https://doi.org/10.1016/j.foodchem.2008.06.014 doi: 10.1016/j.foodchem.2008.06.014
|
| [7] |
Wille JJ, Berhow MA (2011) Bioactives derived from ripe corn tassels: A possible new natural skin whitener, 4-hydroxy-1-oxindole-3-acetic acid. Curr Bioact Compd 7: 126–134. https://doi.org/10.2174/157340711796011115 doi: 10.2174/157340711796011115
|
| [8] |
Žilić S, Vančetović J, Janković M, et al. (2014) Chemical composition, bioactive compounds, antioxidant capacity and stability of floral maize (Zea mays L.) pollen. J Funct Foods 10: 65–74. https://doi.org/10.1016/j.jff.2014.05.007 doi: 10.1016/j.jff.2014.05.007
|
| [9] |
Al-Khayri JM, Yüksel AK, Yüksel M, et al. (2022) Phenolic profile and antioxidant, anticholinergic, and antibacterial properties of corn tassel. Plants 11: 1899. https://doi.org/10.3390/plants11151899 doi: 10.3390/plants11151899
|
| [10] |
Wang L, Yu Y, Fang M, et al. (2014) Antioxidant and antigenotoxic activity of bioactive extracts from corn tassel. J Huazhong Univ Sci Technol Med Sci 34: 131–136. https://doi.org/10.1007/s11596-014-1244-x doi: 10.1007/s11596-014-1244-x
|
| [11] |
Elsayed N, Marrez DA, Ali MA, et al. (2022) Phenolic profiling and in-vitro bioactivities of corn (Zea mays L.) tassel extracts by combining enzyme-assisted extraction. Foods 11: 2145. https://doi.org/10.3390/foods11142145 doi: 10.3390/foods11142145
|
| [12] |
Yaman B (2022) Medical physiological perspective to biochemical assays and GC-MS results of corn tassel. Int J Second Metab 9: 513–524. https://doi.org/10.21448/ijsm.1148489 doi: 10.21448/ijsm.1148489
|
| [13] |
Duangpapeng P, Suriharn K, Lertrat K, et al. (2023) Pigmentation and genotype effects, phenotypic stability for anthocyanins, phenolic compounds and antioxidant activity in the corn tassel. Agrivita 45: 110–123. http://doi.org/10.17503/agrivita.v45i1.3680 doi: 10.17503/agrivita.v45i1.3680
|
| [14] |
Bujang JS, Zakaria MH, Ramaiya SD (2021) Chemical constituents and phytochemical properties of floral maize pollen. PLoS One 16: e0247327. http://doi.org/10.1371/journal.pone.0247327 doi: 10.1371/journal.pone.0247327
|
| [15] |
Duangpapeng P, Ketthaisong D, Lomthaisong K, et al. (2018) Corn tassel: A new source of phytochemicals and antioxidant potential for value-added product development in the agro-industry. Agronomy 8: 242. https://doi.org/10.3390/agronomy8110242 doi: 10.3390/agronomy8110242
|
| [16] |
Duangpapeng P, Lertrat K, Lomthaisong K, et al. (2019) Variability in anthocyanins, phenolic compounds and antioxidant capacity in the tassels of collected waxy corn germplasm. Agronomy 9: 158. https://doi.org/10.3390/agronomy9030158 doi: 10.3390/agronomy9030158
|
| [17] |
Tamprasit K, Weerapreeyakul N, Sutthanut K, et al. (2019) Harvest age effect on phytochemical content of white and black glutinous rice cultivars. Molecules 24: 4432. https://doi.org/10.3390/molecules24244432 doi: 10.3390/molecules24244432
|
| [18] |
Lee J, Durst RW, Wrolstad RE, et al. (2005) Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the pH differential method: Collaborative study. J AOAC Int 88: 1269–1278. https://doi.org/10.1093/jaoac/88.5.1269 doi: 10.1093/jaoac/88.5.1269
|
| [19] |
Yongpradoem P, Weerapreeyakul N (2020) Evaluation of antioxidant activity and inhibition of tyrosinase activity of Raphanus sativus var. caudatus Alef extract. Walailak J Sci Technol 17: 838–850. https://doi.org/10.48048/wjst.2020.5541 doi: 10.48048/wjst.2020.5541
|
| [20] | Gomez KA, Gomez AA (1984) Statistical Procedures for Agricultural Research. 2nd ed, John Wiley and Sons, Singapore. |
| [21] |
Thitilertdecha N (2022) Storage effect on phenolic compounds and antioxidant activity of Nephelium lappaceum L. extract. Cosmetics 9: 33. https://doi.org/10.3390/cosmetics9020033 doi: 10.3390/cosmetics9020033
|
| [22] |
Enaru B, Drețcanu G, Pop TD, et al. (2021) Anthocyanins: Factors affecting their stability and degradation. Antioxidants 10: 1967. https://doi.org/10.3390/antiox10121967 doi: 10.3390/antiox10121967
|
| [23] |
Martini D, Taddei F, Ciccoritti R, et al. (2015) Variation of total antioxidant activity and of phenolic acid, total phenolics and yellow coloured pigments in durum wheat (Triticum turgidum L. var. durum) as a function of genotype, crop year and growing area. J Cereal Sci 65: 175–185. https://doi.org/10.1016/j.jcs.2015.06.012 doi: 10.1016/j.jcs.2015.06.012
|
| [24] |
Fonseca AE, Westgate ME (2005) Relationship between desiccation and viability of maize pollen. Field Crops Res 94: 114–125. https://doi.org/10.1016/j.fcr.2004.12.001 doi: 10.1016/j.fcr.2004.12.001
|
| [25] |
van Bilsen DGJL, Hoekstra FA, Crowe LM, et al. (1994) Altered phase behavior in membranes of aging dry pollen may cause imbibitional leakage. Plant Physiol 104: 1193–1199. https://doi.org/10.1104/pp.104.4.1193 doi: 10.1104/pp.104.4.1193
|
| [26] |
Del-Toro-Sánchez CL, Gutiérrez-Lomelí M, Lugo-Cervantes E, et al. (2015) Storage effect on phenols and on the antioxidant activity of extracts from Anemopsis californica and inhibition of elastase enzyme. J Chem 2015: 602136. https://doi.org/10.1155/2015/602136 doi: 10.1155/2015/602136
|
| [27] |
Moldovan B, Popa A, David L (2016) Effects of storage temperature on the total phenolic content of Cornelian Cherry (Cornus mas L.) fruits extracts. J Appl Bot Food Qual 89: 208–211. https://doi.org/10.5073/JABFQ.2016.089.026 doi: 10.5073/JABFQ.2016.089.026
|
| [28] |
Vicente SJV, Queiroz YS, Gotlieb SLD, et al. (2014) Stability of phenolic compounds and antioxidant capacity of regular and decaffeinated coffees. Braz Arch Biol Technol 57: 110–118. https://doi.org/10.1590/S1516-89132014000100016 doi: 10.1590/S1516-89132014000100016
|
| [29] |
Hernández-Herrero JA, Frutos MJ (2011) Degradation kinetics of pigment, colour and stability of the antioxidant capacity in juice model systems from six anthocyanin sources. In J Food Sci Technol 46: 2550–2557. https://doi.org/10.1111/j.1365-2621.2011.02780.x doi: 10.1111/j.1365-2621.2011.02780.x
|
| [30] |
Deng LZ, Xiong CH, Pei YP, et al. (2022) Effects of various storage conditions on total phenolic, carotenoids, antioxidant capacity, and color of dried apricots. Food Control 136: 108846. https://doi.org/10.1016/j.foodcont.2022.108846 doi: 10.1016/j.foodcont.2022.108846
|
| [31] |
Deng M, Deng Y, Dong L, et al. (2018) Effect of storage conditions on phenolic profiles and antioxidant activity of litchi pericarp. Molecules 23: 2276. https://doi.org/10.3390/molecules23092276 doi: 10.3390/molecules23092276
|
| [32] |
Khoo HE, Azlan A, Tang ST, et al. (2017) Anthocyanidins and anthocyanins: colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr Res 61: 1361779. https://doi.org/10.1080/16546628.2017.1361779 doi: 10.1080/16546628.2017.1361779
|
| [33] |
Irani NG, Grotewold E (2005) Light-induced morphological alteration in anthocyanin-accumulating vacuoles of maize cells. BMC Plant Biol 5: 7. https://doi.org/10.1186/1471-2229-5-7 doi: 10.1186/1471-2229-5-7
|
| [34] |
Cone KC, Cocciolone SM, Burr FA, et al. (1993) Maize anthocyanin regulatory gene pl is a duplicate of c1 that functions in the plant. Plant Cell 5: 1795. https://doi.org/10.1105/tpc.5.12.1795 doi: 10.1105/tpc.5.12.1795
|
| [35] |
Kim S, Hwang G, Lee S, (2017) High ambient temperature represses anthocyanin biosynthesis through degradation of HY5. Front Plant Sci 8: 1787. https://doi.org/10.3389/fpls.2017.01787 doi: 10.3389/fpls.2017.01787
|
| [36] |
Hellström J, Mattila P, Karjalainen R (2013) Stability of anthocyanins in berry juices stored at different temperatures. J Food Compos Anal 31: 12–19. https://doi.org/10.1016/j.jfca.2013.02.010 doi: 10.1016/j.jfca.2013.02.010
|
| [37] |
Muche BM, Speers RA, Rupasinghe HPV (2018) Storage temperature impacts on anthocyanins degradation, color changes and haze development in juice of "Merlot" and "Ruby" grapes (Vitis vinifera). Front Nutr 5: 100. https://doi.org/10.3389/fnut.2018.00100 doi: 10.3389/fnut.2018.00100
|
| [38] |
Fracassetti D, Del Bo C, Simonetti P, et al. (2013) Effect of time and storage temperature on anthocyanin decay and antioxidant activity in wild blueberry (Vaccinium angustifolium) powder. J Agric Food Chem 61: 2999–3005. https://doi.org/10.1021/jf3048884 doi: 10.1021/jf3048884
|
| [39] |
Oliveira KG, Queiroz VAV, Carlos LA, et al. (2017) Effect of the storage time and temperature on phenolic compounds of sorghum grain and flour. Food Chem 216: 390–398. https://doi.org/10.1016/j.foodchem.2016.08.047 doi: 10.1016/j.foodchem.2016.08.047
|
| [40] |
Harakotr B, Suriharn B, Tangwongchai R, et al. (2014) Anthocyanin, phenolics and antioxidant activity changes in purple waxy corn as affected by traditional cooking. Food Chem 164: 510–517. https://doi.org/10.1016/j.foodchem.2014.05.069 doi: 10.1016/j.foodchem.2014.05.069
|
| [41] |
Shiekh KA, Luanglaor T, Hanprerakriengkrai N, et al. (2023) Antioxidants and quality changes of thermally processed purple corn (Zea mays L.) milk fortified with low sucrose content during cold storage. Foods 12: 277. https://doi.org/10.3390/foods12020277 doi: 10.3390/foods12020277
|
| [42] |
Charmongkolpradit S, Somboon T, Phatchana R, et al. (2021) Influence of drying temperature on anthocyanin and moisture contents in purple waxy corn kernel using a tunnel dryer. Case Stud Therm Eng 25: 100886. https://doi.org/10.1016/j.csite.2021.100886 doi: 10.1016/j.csite.2021.100886
|
| [43] |
Ursu MS, Aprodu I, Milea SA, et al. (2020) Thermal degradation kinetics of anthocyanins extracted from purple maize flour extract and the effect of heating on selected biological functionality. Foods 9: 1593. https://doi.org/10.3390/foods9111593 doi: 10.3390/foods9111593
|
| [44] | Olugbami JO, Gbadegesin MA, Odunola OA (2014) In vitro evaluation of the antioxidant potential, phenolic and flavonoid contents of the stem bark ethanol extract of Anogeissus leiocarpus. Afr J Med Med Sci 43(Suppl 1): 101–109. |
| [45] |
Lin YS, Huang WY, Ho PY, et al. (2020) Effects of storage time and temperature on antioxidants in juice from Momordica charantia L. and Momordica charantia L. var. abbreviata Ser. Molecules 25: 3614. https://doi.org/10.3390/molecules25163614 doi: 10.3390/molecules25163614
|
| [46] |
Ratha J, Yongram C, Panyatip P, et al, (2023) Polyphenol and tryptophan contents of purple corn (Zea mays L.) variety KND and butterfly pea (Clitoria ternatea) aqueous extracts: Insights into phytochemical profiles with antioxidant activities and PCA analysis. Plants 12: 603. https://doi.org/10.3390/plants12030603 doi: 10.3390/plants12030603
|
| [47] |
Bae HH, Yi G, Go YS, et al. (2021) Measuring antioxidant activity in yellow corn (Zea mays L.) inbreds from three different geographic regions. Appl Biol Chem 64: 56. https://doi.org/10.1186/s13765-021-00629-y doi: 10.1186/s13765-021-00629-y
|
| [48] |
Boateng ID, Kumar R, Daubert CR, et al. (2023) Sonoprocessing improves phenolics profile, antioxidant capacity, structure, and product qualities of purple corn pericarp extract. Ultrason Sonochem 95: 106418. https://doi.org/10.1016/j.ultsonch.2023.106418 doi: 10.1016/j.ultsonch.2023.106418
|

Figures(2) / Tables(2)

Prakasit Duangpapeng, Abil Dermail, Khundej Suriharn. Phenolics, anthocyanins, and antioxidant capacity in the tassels of purple waxy corn: Effects of temperature and time during storage[J]. AIMS Agriculture and Food, 2024, 9(1): 69-83. doi: 10.3934/agrfood.2024005







 DownLoad:
DownLoad: