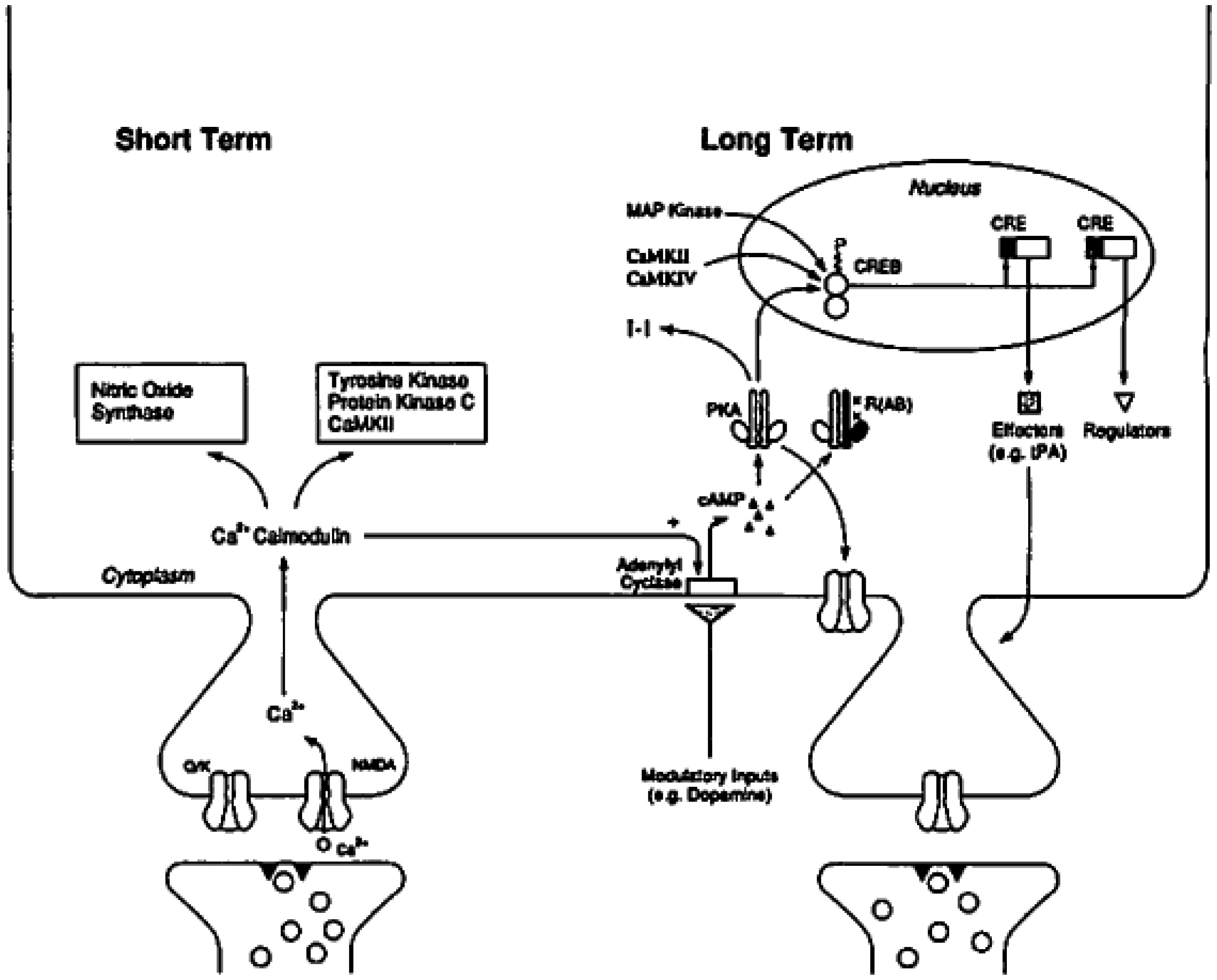
I recount some landmark discoveries that initially confirmed the cyclic AMP response element-binding (CREB) protein-memory consolidation and allocation linkages. This work constitutes one of the successes of the field of Molecular and Cellular Cognition (MCC) but is also of interest to philosophers of neuroscience. Two approaches, “mechanism” and “ruthless reductionism”, claim to account for this case, yet these accounts differ in one crucial way. I explain this difference and argue that both the experiment designs and discussions of these discoveries by MCC scientists better fit the ruthless reductionist's account. This conclusion leads to further philosophical discussion about how discoveries in cellular/molecular neurobiology integrate with systems neuroscience findings.
Citation: John Bickle. The first two decades of CREB-memory research: data for philosophy of neuroscience[J]. AIMS Neuroscience, 2021, 8(3): 322-339. doi: 10.3934/Neuroscience.2021017
I recount some landmark discoveries that initially confirmed the cyclic AMP response element-binding (CREB) protein-memory consolidation and allocation linkages. This work constitutes one of the successes of the field of Molecular and Cellular Cognition (MCC) but is also of interest to philosophers of neuroscience. Two approaches, “mechanism” and “ruthless reductionism”, claim to account for this case, yet these accounts differ in one crucial way. I explain this difference and argue that both the experiment designs and discussions of these discoveries by MCC scientists better fit the ruthless reductionist's account. This conclusion leads to further philosophical discussion about how discoveries in cellular/molecular neurobiology integrate with systems neuroscience findings.

| [1] | Dudai Y (2004) Memory from A to Z: Keywords, Concepts, and Beyond New York: Oxford University Press. |
| [2] |
Davis HR, Squire LR (1984) Protein synthesis and memory: A review. Psychol Bull 96: 518-559. doi: 10.1037/0033-2909.96.3.518

|
| [3] |
Duncan CP (1949) The retroactive effect of electroshock on learning. J Comp Physiol Psychol 42: 32-44. doi: 10.1037/h0058173
|
| [4] |
Dash PK, Hochner B, Kandel ER (1990) Injection of the cAMP-responsive element into the nucleus of Aplysia sensory neurons blocks long-term facilitation. Nature 345: 718-721. doi: 10.1038/345718a0
|
| [5] |
Sheng M, Thompson MA, Greenberg ME (1991) CREB: A Ca2+-regulated transcription factor phosphorylated by calmodulin-dependent kinases. Science 252: 1427-1430. doi: 10.1126/science.1646483
|
| [6] |
Silva AJ, Stevens CF, Tonegawa S, et al. (1992) Deficient hippocampal long-term potentiation in alpha-calcium-calmodulin kinase II mutant mice. Science 257: 201-206. doi: 10.1126/science.1378648
|
| [7] |
Silva AJ, Paylor R, Wehner JM, et al. (1992b) Impaired spatial learning in alpha-calcium-calmodulin kinase II mutant mice. Science 257: 206-211. doi: 10.1126/science.1321493
|
| [8] | Silva AJ, Landreth A, Bickle J (2014) Engineering the Next Revolution in Neuroscience New York: Oxford University Press. |
| [9] |
Hummler E, Cole TJ, Blendy JA, et al. (1994) Targeted mutation of the CREB gene: Compensation within the CREB/ATF family of transcription factors. Proc Natl Acad Sci USA 91: 5647-5651. doi: 10.1073/pnas.91.12.5647
|
| [10] |
Bourtchouladze R, Frenguelli B, Blendy J, et al. (1994) Deficient long-term memory in mice with a targeted mutation of the cAMP-responsive element-binding protein. Cell 79: 59-68. doi: 10.1016/0092-8674(94)90400-6
|
| [11] |
Bickle J (2003) Philosophy and Neuroscience: A ruthlessly reductive approach Dordrecht: Springer. doi: 10.1007/978-94-010-0237-0
|
| [12] |
Alberini CM (2009) Transcription factors in long-term memory and synaptic plasticity. Physiol Rev 89: 121-145. doi: 10.1152/physrev.00017.2008
|
| [13] |
Abel T, Nguyen PV, Barad M, et al. (1997) Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus-based long-term memory. Cell 88: 615-626. doi: 10.1016/S0092-8674(00)81904-2
|
| [14] | Purves D, Augustine GL, Fitzpatrick D, et al. (2017) Neuroscience New York: Oxford University Press. |
| [15] |
Alberini CM, Kandel ER (2015) The regulation of transcription in memory consolidation. Cold Spring Harb Perspect Biol 7: a021741. doi: 10.1101/cshperspect.a021741
|
| [16] |
Josselyn SA, Shi C, Carlezon WA, et al. (2001) Long-term memory is facilitated by cAMP responsive element-binding protein overexpression in the amygdala. J Neurosci 21: 2404-2412. doi: 10.1523/JNEUROSCI.21-07-02404.2001
|
| [17] |
Han JH, Kushner SA, Yiu AP, et al. (2007) Neuronal competition and selection during memory formation. Science 316: 457-460. doi: 10.1126/science.1139438
|
| [18] |
Guzowski JF, McNaughton BI, Barnes CA, et al. (1999) Environment-specific expression of the immediate-early gene Arc in hippocampal neuronal assemblies. Nat Neurosci 2: 1120-1124. doi: 10.1038/16046
|
| [19] |
Zhou Y, Won J, Karlsson MG, et al. (2009) CREB regulates excitability and the allocation of memory to subsets of neurons in the amygdala. Nat Neurosci 12: 1438-1443. doi: 10.1038/nn.2405
|
| [20] |
Han JH, Kushner SA, You AP, et al. (2009) Selective erasure of a fear memory. Science 323: 1492-1496. doi: 10.1126/science.1164139
|
| [21] | Bickle J, Mandik P, Landreth A (2019) Philosophy of neuroscience. The Stanford Encyclopedia of Philosophy Available from: https://plato.stanford.edu/archives/fall2019)/entries/neuroscience/. |
| [22] | Bechtel W (2008) Mental Mechanisms: Philosophical perspectives on cognitive neuroscience London: Routledge. |
| [23] | Bechtel W (2009) Molecules systems, and behavior: Another view of memory consolidation. The Oxford Handbook of Philosophy and Neuroscience New York: Oxford University Press, 13-40. |
| [24] |
Craver CF (2007) Explaining the Brain New York: Oxford University Press. doi: 10.1093/acprof:oso/9780199299317.001.0001
|
| [25] |
Craver CF, Darden L (2013) In Search of Mechanisms: Discoveries Across the Life Sciences Chicago: University of Chicago Press. doi: 10.7208/chicago/9780226039824.001.0001
|
| [26] |
Bickle J (2006) Reducing mind to molecular pathways: Explicating the reductionism implicit in current cellular and molecular neuroscience. Synthese 151: 411-434. doi: 10.1007/s11229-006-9015-2
|
| [27] | Bickle J (2020) Laser lights and designer drugs: New techniques for descending levels of mechanisms “in a single bound”? TopiCS 12: 1241-1256. |
| [28] |
Barwich AS (2020) Smellosophy: What the nose tells the mind Cambridge, MA: Harvard University Press. doi: 10.4159/9780674245426
|
Figures(2)

John Bickle. The first two decades of CREB-memory research: data for philosophy of neuroscience[J]. AIMS Neuroscience, 2021, 8(3): 322-339. doi: 10.3934/Neuroscience.2021017







 DownLoad:
DownLoad: