Citation: Mônica H. M. Nascimento, Milena T. Pelegrino, Joana C. Pieretti, Amedea B. Seabra. How can nitric oxide help osteogenesis?[J]. AIMS Molecular Science, 2020, 7(1): 29-48. doi: 10.3934/molsci.2020003

| [1] |
Feng X, Mcdonald JM (2011) Disorders of Bone Remodeling. Annu Rev Pathol 6: 121-145. doi: 10.1146/annurev-pathol-011110-130203

|
| [2] |
Van'T Hof RJ, Ralston SH (2001) Nitric oxide and bone. Immunology 103: 255-261. doi: 10.1046/j.1365-2567.2001.01261.x
|
| [3] |
Parfitt AM (1987) Bone remodeling and bone loss: Understanding the pathophysiology of osteoporosis. Clin Obstet Gynecol 30: 789-811. doi: 10.1097/00003081-198712000-00004
|
| [4] |
Diwan AD, Wang MX, Jang D, et al. (2000) Nitric Oxide Modulates Fracture Healing. J Bone Miner Res 15: 342-351. doi: 10.1359/jbmr.2000.15.2.342
|
| [5] |
Pandya CD, Lee B, Toque HA, et al. (2019) Age-Dependent Oxidative Stress Elevates Arginase 1 and Uncoupled Nitric Oxide Synthesis in Skeletal Muscle of Aged Mice. Oxid Med Cell Longev 2019: 1-9. doi: 10.1155/2019/1704650
|
| [6] |
Aguirre J, Buttery LDK, O'Shaughnessy M, et al. (2001) Endothelial nitric oxide synthase gene-deficient mice demonstrate marked retardation in postnatal bone formation, reduced bone volume, and defects in osteoblast maturation and activity. Am J Pathol 158: 247-257. doi: 10.1016/S0002-9440(10)63963-6
|
| [7] |
Ignarro LJ (1999) Nitric Oxide: A Unique Endogenous Signaling Molecule in Vascular Biology (Nobel Lecture). Angew Chemie Int Ed 38: 1882-1892. doi: 10.1002/(SICI)1521-3773(19990712)38:13/14<1882::AID-ANIE1882>3.0.CO;2-V
|
| [8] |
Koshland D (1992) The molecule of the year. Science 258: 1861. doi: 10.1126/science.1470903
|
| [9] |
MacMicking J, Xie Q, Nathan C (1997) Nitric oxide and macrophage function. Annu Rev Immunol 15: 323-350. doi: 10.1146/annurev.immunol.15.1.323
|
| [10] |
Culotta E, Koshland DE (1992) NO news is good news. Science 258: 1862-1865. doi: 10.1126/science.1361684
|
| [11] |
Tousoulis D, Kampoli A, Tentolouris C, et al. (2012) The Role of Nitric Oxide on Endothelial Function. Curr Vasc Pharmacol 10: 4-18. doi: 10.2174/157016112798829760
|
| [12] |
Basudhar D, Cheng RC, Bharadwaj G, et al. (2015) Chemotherapeutic potential of diazeniumdiolate-based aspirin prodrugs in breast cancer. Free Radic Biol Med 83: 101-114. doi: 10.1016/j.freeradbiomed.2015.01.029
|
| [13] |
Seabra AB, Duran N (2017) Nanoparticulated Nitric Oxide Donors and their Biomedical Applications. Mini Rev Med Chem 17: 216-223. doi: 10.2174/1389557516666160808124624
|
| [14] |
Rolim WR, Pieretti JC, Renó DLS, et al. (2019) Antimicrobial Activity and Cytotoxicity to Tumor Cells of Nitric Oxide Donor and Silver Nanoparticles Containing PVA/PEG Films for Topical Applications. ACS Appl Mater Interfaces 11: 6589-6604. doi: 10.1021/acsami.8b19021
|
| [15] |
Kalyanaraman H, Schall N, Pilz RB (2018) Nitric oxide and cyclic GMP functions in bone. Nitric Oxide Biol Chem 76: 62-70. doi: 10.1016/j.niox.2018.03.007
|
| [16] |
Yang C, Jeong S, Ku S, et al. (2018) Use of gasotransmitters for the controlled release of polymer-based nitric oxide carriers in medical applications. J Control Release 279: 157-170. doi: 10.1016/j.jconrel.2018.04.025
|
| [17] |
Seabra AB, Pelegrino MT, Haddad PS (2016) Can Nitric Oxide Overcome Bacterial Resistance to Antibiotics? Antibiotic Resistance Elsevier, 187-204. doi: 10.1016/B978-0-12-803642-6.00009-5
|
| [18] |
Andrew PJ, Mayer B (1999) Enzymatic function of nitric oxide synthases. Cardiovasc Res 43: 521-531. doi: 10.1016/S0008-6363(99)00115-7
|
| [19] |
Forstermann U, Sessa WC (2012) Nitric oxide synthases: regulation and function. Eur Hear Journa 33: 829-837. doi: 10.1093/eurheartj/ehr304
|
| [20] |
Hutchinson PJA, Palmer RMJ, Moncada S (1987) Comparative pharmacology of EDRF and nitric oxide on vascular strips. Eur J Pharmacol 141: 445-451. doi: 10.1016/0014-2999(87)90563-2
|
| [21] |
Pelegrino M, de Araujo Lima B, do Nascimento M, et al. (2018) Biocompatible and Antibacterial Nitric Oxide-Releasing Pluronic F-127/Chitosan Hydrogel for Topical Applications. Polymers (Basel) 10: 452. doi: 10.3390/polym10040452
|
| [22] |
Pelegrino MT, Silva LC, Watashi CM, et al. (2017) Nitric oxide-releasing nanoparticles: synthesis, characterization, and cytotoxicity to tumorigenic cells. J Nanoparticle Res 19: 57. doi: 10.1007/s11051-017-3747-4
|
| [23] |
Ferraz LS, Watashi CM, Colturato-Kido C, et al. (2018) Antitumor Potential of S-Nitrosothiol-Containing Polymeric Nanoparticles against Melanoma. Mol Pharm 15: 1160-1168. doi: 10.1021/acs.molpharmaceut.7b01001
|
| [24] |
Aranda E, Lopez-Pedrera C, De La Haba-Rodriguez JR, et al. (2012) Nitric Oxide and Cancer: The Emerging Role of S-Nitrosylation. Curr Mol Med 12: 50-67. doi: 10.2174/156652412798376099
|
| [25] |
González R, Molina-Ruiz FJ, Bárcena JA, et al. (2018) Regulation of Cell Survival, Apoptosis, and Epithelial-to-Mesenchymal Transition by Nitric Oxide-Dependent Post-Translational Modifications. Antioxidants Redox Signal 29: 1312-1332. doi: 10.1089/ars.2017.7072
|
| [26] |
Kollau A, Russwurm M, Neubauer A, et al. (2016) Scavenging of nitric oxide by hemoglobin in the tunica media of porcine coronary arteries. Nitric Oxide 54: 8-14. doi: 10.1016/j.niox.2016.01.005
|
| [27] |
Larry KK, Raymond WN, Keith MD, et al. (1996) “NONOates” (1-Substituted Diazen-1-ium-1,2-diolates) as Nitric Oxide Donors: Convenient Nitric Oxide Dosage Forms. Methods Enzymol 268: 281-293. doi: 10.1016/S0076-6879(96)68030-6
|
| [28] |
Wang PG, Xian M, Tang X, et al. (2002) Nitric oxide donors: Chemical activities and biological applications. Chem Rev 102: 1091-1134. doi: 10.1021/cr000040l
|
| [29] |
Liang H, Nacharaju P, Friedman A, et al. (2015) Nitric oxide generating/releasing materials. Futur Sci OA 1: FSO54. doi: 10.4155/fso.15.54
|
| [30] |
da Silva RS, de Lima RG, de Paula Machado S (2015) Design, reactivity, and biological activity of ruthenium nitrosyl complexes. Adv Inorg Chem 67: 265-294. doi: 10.1016/bs.adioch.2014.11.001
|
| [31] |
Seabra AB, De Souza GFP, Da Rocha LL, et al. (2004) S-Nitrosoglutathione incorporated in poly(ethylene glycol) matrix: Potential use for topical nitric oxide delivery. Nitric Oxide Biol Chem 11: 263-272. doi: 10.1016/j.niox.2004.09.005
|
| [32] |
Seabra AB, Fitzpatrick A, Paul J, et al. (2004) Topically applied S-nitrosothiol-containing hydrogels as experimental and pharmacological nitric oxide donors in human skin. Br J Dermatol 151: 977-983. doi: 10.1111/j.1365-2133.2004.06213.x
|
| [33] |
Oliveira HC, Gomes BCR, Pelegrino MT, et al. (2016) Nitric oxide-releasing chitosan nanoparticles alleviate the effects of salt stress in maize plants. Nitric Oxide Biol Chem 61: 10-19. doi: 10.1016/j.niox.2016.09.010
|
| [34] |
Heinonen I, Boushel R, Hellsten Y, et al. (2018) Regulation of bone blood flow in humans: The role of nitric oxide, prostaglandins, and adenosine. Scand J Med Sci Sports 28: 1552-1558. doi: 10.1111/sms.13064
|
| [35] |
Borys J, Maciejczyk M, Antonowicz B, et al. (2019) Glutathione Metabolism, Mitochondria Activity, and Nitrosative Stress in Patients Treated for Mandible Fractures. J Clin Med 8: 127. doi: 10.3390/jcm8010127
|
| [36] | Sela JJ, Bab IA (2012) Healing of Bone Fracture: General Concepts. Principles of Bone Regeneration Boston, MA: Springer US, 1-8. |
| [37] |
Helfrich MH, Evans DE, Grabowski PS, et al. (1997) Expression of Nitric Oxide Synthase Isoforms in Bone and Bone Cell Cultures. J Bone Miner Res 12: 1108-1115. doi: 10.1359/jbmr.1997.12.7.1108
|
| [38] |
Basso N, Heersche JNM (2006) Effects of hind limb unloading and reloading on nitric oxide synthase expression and apoptosis of osteocytes and chondrocytes. Bone 39: 807-814. doi: 10.1016/j.bone.2006.04.014
|
| [39] |
Bakker AD, Huesa C, Hughes A, et al. (2013) Endothelial Nitric Oxide Synthase is Not Essential for Nitric Oxide Production by Osteoblasts Subjected to Fluid Shear Stress In Vitro. Calcif Tissue Int 92: 228-239. doi: 10.1007/s00223-012-9670-x
|
| [40] |
Grassi F, Fan X, Rahnert J, et al. (2006) Bone Re/Modeling Is More Dynamic in the Endothelial Nitric Oxide Synthase (−/−) Mouse. Endocrinology 147: 4392-4399. doi: 10.1210/en.2006-0334
|
| [41] |
MacPherson H, Noble BS, Ralston SH (1999) Expression and functional role of nitric oxide synthase isoforms in human osteoblast-like cells. Bone 24: 179-185. doi: 10.1016/S8756-3282(98)00173-2
|
| [42] |
Zheng H, Yu X, Collin-Osdoby P, et al. (2006) RANKL Stimulates Inducible Nitric-oxide Synthase Expression and Nitric Oxide Production in Developing Osteoclasts. J Biol Chem 281: 15809-15820. doi: 10.1074/jbc.M513225200
|
| [43] |
Samuels A, Perry MJ, Gibson RL, et al. (2001) Role of endothelial nitric oxide synthase in estrogen-induced osteogenesis. Bone 29: 24-29. doi: 10.1016/S8756-3282(01)00471-9
|
| [44] |
Van'T Hof RJ, Macphee J, Libouban H, et al. (2004) Regulation of bone mass and bone turnover by neuronal nitric oxide synthase. Endocrinology 145: 5068-5074. doi: 10.1210/en.2004-0205
|
| [45] |
Kasten TP, Collin-Osdoby P, Patel N, et al. (1994) Potentiation of osteoclast bone-resorption activity by inhibition of nitric oxide synthase. Proc Natl Acad Sci U S A 91: 3569-3573. doi: 10.1073/pnas.91.9.3569
|
| [46] |
Löwik CWGM, Nibbering PH, Van De Ruit M, et al. (1994) Inducible production of nitric oxide in osteoblast-like cells and in fetal mouse bone explants is associated with suppression of osteoclastic bone resorption. J Clin Invest 93: 1465-1472. doi: 10.1172/JCI117124
|
| [47] |
Wimalawansa SJ (2007) Rationale for using nitric oxide donor therapy for prevention of bone loss and treatment of osteoporosis in humans. Ann N Y Acad Sci 1117: 283-297. doi: 10.1196/annals.1402.066
|
| [48] |
Nichols SP, Storm WL, Koh A, et al. (2012) Local delivery of nitric oxide: Targeted delivery of therapeutics to bone and connective tissues. Adv Drug Deliv Rev 64: 1177-1188. doi: 10.1016/j.addr.2012.03.002
|
| [49] |
Rosselli M, Imthurn B, Keller PJ, et al. (1995) Circulating nitric oxide (nitrite/nitrate) levels in postmenopausal women substituted with 17β-estradiol and norethisterone acetate: A two-year follow-up study. Hypertension 25: 848-853. doi: 10.1161/01.HYP.25.4.848
|
| [50] |
Ralston SH, Ho L, Helfrich MH, et al. (1995) Nitric Oxide?: A Cytokine-Induced Regulator of. J Bone Miner Res 10: 1040-1049. doi: 10.1002/jbmr.5650100708
|
| [51] |
Ralston SH, Grabowski PS (1996) Mechanisms of cytokine induced bone resorption: Role of nitric oxide, cyclic guanosine monophosphate, and prostaglandins. Bone 19: 29-33. doi: 10.1016/8756-3282(96)00101-9
|
| [52] |
Wimalawansa SJ (2010) Nitric oxide and bone. Ann NY Acad Sci 1192: 394-406. doi: 10.1111/j.1749-6632.2009.05230.x
|
| [53] |
Turner CH, Owan I, Jacob DS, et al. (1997) Effects of nitric oxide synthase inhibitors on bone formation in rats. Bone 21: 487-490. doi: 10.1016/S8756-3282(97)00202-0
|
| [54] |
Mancini L, Moradi-Bidhendi N, Becherini L, et al. (2000) The biphasic effects of nitric oxide in primary rat osteoblasts are cGMP dependent. Biochem Biophys Res Commun 274: 477-481. doi: 10.1006/bbrc.2000.3164
|
| [55] |
Chambers TJ, Fox S, Jagger CJ, et al. (1999) The role of prostaglandins and nitric oxide in the response of bone to mechanical forces. Osteoarthritis Cartilage 7: 422-423. doi: 10.1053/joca.1998.0231
|
| [56] |
Chow JWM, Fox SW, Lean JM, et al. (1998) Role of nitric oxide and prostaglandins in mechanically induced bone formation. J Bone Miner Res 13: 1039-1044. doi: 10.1359/jbmr.1998.13.6.1039
|
| [57] |
Wimalawansa SJ, De Marco G, Gangula P, et al. (1996) Nitric oxide donor alleviates ovariectomy-induced bone loss. Bone 18: 301-304. doi: 10.1016/8756-3282(96)00005-1
|
| [58] |
Park YG, Kim KW, Song KH, et al. (2009) Combinatory responses of proinflamamtory cytokines on nitric oxide-mediated function in mouse calvarial osteoblasts. Cell Biol Int 33: 92-99. doi: 10.1016/j.cellbi.2008.09.012
|
| [59] |
Cuzzocrea S, Mazzon E, Dugo L, et al. (2003) Inducible Nitric Oxide Synthase Mediates Bone Loss in Ovariectomized Mice. Endocrinology 144: 1098-1107. doi: 10.1210/en.2002-220597
|
| [60] |
Robling AG, Castillo AB, Turner CH (2006) Biomechanical and Molecular Regulation of Bone Remodeling. Annu Rev Biomed Eng 8: 455-498. doi: 10.1146/annurev.bioeng.8.061505.095721
|
| [61] |
Mcallister TN, Du T, Frangos JA (2000) Fluid Shear Stress Stimulates Prostaglandin and Nitric Oxide Release in Bone Marrow-Derived. Biochem Biophys Res Commun 270: 643-648. doi: 10.1006/bbrc.2000.2467
|
| [62] |
Abnosi MH, Pari S (2019) Exogenous nitric oxide induced early mineralization in rat bone marrow mesenchymal stem cells via activation of alkaline phosphatase. Iran Biomed J 23: 142-152. doi: 10.29252/ibj.23.2.142
|
| [63] |
Ocarino NM, Boeloni JN, Goes AM, et al. (2008) Nitric Oxide Osteogenic differentiation of mesenchymal stem cells from osteopenic rats subjected to physical activity with and without nitric oxide synthase inhibition. Nitric Oxide 19: 320-325. doi: 10.1016/j.niox.2008.08.004
|
| [64] |
Lee JS, Lee HJ, Lee JW, et al. (2018) Osteogenic Effect of Inducible Nitric Oxide Synthase (iNOS)-Loaded Mineralized Nanoparticles on Embryonic Stem Cells. Cell Physiol Biochem 51: 746-762. doi: 10.1159/000495330
|
| [65] |
Fuseler JW, Valarmathi MT (2016) Nitric Oxide Modulates Postnatal Bone Marrow-Derived Mesenchymal. Front Cell Dev Biol 4: 1-20. doi: 10.3389/fcell.2016.00133
|
| [66] |
Takahashi TA, Johnson KM (2015) Menopause. Med Clin North Am 99: 521-534. doi: 10.1016/j.mcna.2015.01.006
|
| [67] |
Daripa M, Paula FJA, Rufino ACB, et al. (2004) Impact of congenital calcitonin deficiency due to dysgenetic hypothyroidism on bone mineral density. Brazilian J Med Biol Res 37: 61-68. doi: 10.1590/S0100-879X2004000100009
|
| [68] |
Wimalawansa SJ (2000) Restoration of ovariectomy-induced osteopenia by nitroglycerin. Calcif Tissue Int 66: 56-60. doi: 10.1007/s002230050011
|
| [69] |
Wimalawansa SJ (2000) Nitroglycerin therapy is as efficacious as standard estrogen replacement therapy (premarin) in prevention of oophorectomy-induced bone loss: A human pilot clinical study. J Bone Miner Res 15: 2240-2244. doi: 10.1359/jbmr.2000.15.11.2240
|
| [70] |
Visser JJ, Hoekman K (1994) Arginine supplementation in the prevention and treatment of osteoporosis. Med Hypotheses 43: 339-342. doi: 10.1016/0306-9877(94)90113-9
|
| [71] |
Fiore CE, Pennisi P, Cutuli VM, et al. (2000) L-arginine prevents bone loss and bone collagen breakdown in cyclosporin A-treated rats. Eur J Pharmacol 408: 323-326. doi: 10.1016/S0014-2999(00)00800-1
|
| [72] |
Veeriah V, Zanniti A, Paone R, et al. (2016) Interleukin-1β, lipocalin 2 and nitric oxide synthase 2 are mechano-responsive mediators of mouse and human endothelial cell-osteoblast crosstalk. Sci Rep 6: 1-14. doi: 10.1038/srep29880
|
| [73] |
Alcaide M, Serrano MC, Pagani R, et al. (2009) Biocompatibility markers for the study of interactions between osteoblasts and composite biomaterials. Biomaterials 30: 45-51. doi: 10.1016/j.biomaterials.2008.09.012
|
| [74] |
Bielemann AM, Marcello-Machado RM, Del Bel Cury AA, et al. (2018) Systematic review of wound healing biomarkers in peri-implant crevicular fluid during osseointegration. Arch Oral Biol 89: 107-128. doi: 10.1016/j.archoralbio.2018.02.013
|
| [75] | Sugiatno E, Samsudin AR, Sosroseno W (2009) Effect of exogenous nitric oxide on the proliferation of a human osteoblast (HOS) cell line induced by hydroxyapatite. J Appl Biomater Biomech 7: 29-33. |
| [76] |
Adhikari U, Rijal NP, Khanal S, et al. (2016) Magnesium incorporated chitosan based scaffolds for tissue engineering applications. Bioact Mater 1: 132-139. doi: 10.1016/j.bioactmat.2016.11.003
|
| [77] |
Herculano RD, Tzu LC, Silva CP, et al. (2011) Nitric oxide release using natural rubber latex as matrix. Mater Res 14: 355-359. doi: 10.1590/S1516-14392011005000055
|
| [78] |
Özmeriç N, Elgün S, Uraz A (2000) Salivary arginase in patients with adult periodontitise. Clin Oral Invest 4: 21-24. doi: 10.1007/s007840050108
|
| [79] |
Leitão RFC, Rocha FAC, Chaves HV, et al. (2004) Locally Applied Isosorbide Decreases Bone Resorption in Experimental Periodontitis in Rats. J Periodontol 75: 1227-1232. doi: 10.1902/jop.2004.75.9.1227
|
| [80] |
Lee SK, Choi HI, Yang YS, et al. (2009) Nitric oxide modulates osteoblastic differentiation with heme oxygenase-1 via the mitogen activated protein kinase and nuclear factor-kappaB pathways in human periodontal ligament cells. Biol Pharm Bull 32: 1328-1334. doi: 10.1248/bpb.32.1328
|
| [81] |
Jönsson D, Nebel D, Bratthall G, et al. (2011) The human periodontal ligament cell: a fibroblast-like cell acting as an immune cell. J Periodontal Res 46: 153-157. doi: 10.1111/j.1600-0765.2010.01331.x
|
| [82] |
de Menezes AMA, de Souza GFP, Gomes AS, et al. (2012) S-Nitrosoglutathione Decreases Inflammation and Bone Resorption in Experimental Periodontitis in Rats. J Periodontol 83: 514-521. doi: 10.1902/jop.2011.110332
|
| [83] | Martins CS, Leitão RFC, Costa DVS, et al. (2016) Topical HPMC/S-Nitrosoglutathione Solution Decreases Inflammation and Bone Resorption in Experimental Periodontal Disease in Rats. PLoS One 11: 1-19. |
| [84] |
Chae HJ, Park RK, Chung HT, et al. (1997) Nitric oxide is a regulator of bone remodelling. J Pharm Pharmacol 49: 897-902. doi: 10.1111/j.2042-7158.1997.tb06132.x
|
| [85] | Lin YJ, Chen CC, Chi NW, et al. (2018) In Situ Self-Assembling Micellar Depots that Can Actively Trap and Passively Release NO with Long-Lasting Activity to Reverse Osteoporosis. Adv Mater 30: 1-6. |
| [86] |
Tai YT, Cherng YG, Chang CC, et al. (2007) Pretreatment with low nitric oxide protects osteoblasts from high nitric oxide-induced apoptotic insults through regulation of c-Jun N-terminal kinase/c-Jun-mediatedBcl-2 gene expression and protein translocation. J Orthop Res 25: 625-635. doi: 10.1002/jor.20365
|
| [87] |
Differ C, Klatte-Schulz F, Bormann N, et al. (2019) Is NO the Answer? The Nitric Oxide Pathway Can Support Bone Morphogenetic Protein 2 Mediated Signaling. Cells 8: 1273. doi: 10.3390/cells8101273
|
| [88] |
Bandara N, Gurusinghe S, Lim SY, et al. (2016) Molecular control of nitric oxide synthesis through eNOS and caveolin-1 interaction regulates osteogenic differentiation of adipose-derived stem cells by modulation of Wnt/β-catenin signaling. Stem Cell Res Ther 7: 1-15. doi: 10.1186/s13287-015-0253-4
|
| [89] |
Chu L, Jiang Y, Hao H, et al. (2008) Nitric oxide enhances Oct-4 expression in bone marrow stem cells and promotes endothelial differentiation. Eur J Pharmacol 591: 59-65. doi: 10.1016/j.ejphar.2008.06.066
|
| [90] | Felka T, Ulrich C, Rolauffs B, et al. (2014) Bone marrow stromal cells Nitric oxide activates signaling by c-Raf, MEK, p-JNK, p38 MAPK and p53 in human mesenchymal stromal cells and inhibits their osteogenic differentiation by blocking expression of Runx2. J stem cell Res Ther 4: 195. |
| [91] |
Jamal SA, Cummings SR, Hawker GA (2004) Isosorbide mononitrate increases bone formation and decreases bone resorption in postmenopausal women: A randomized trial. J Bone Miner Res 19: 1512-1517. doi: 10.1359/JBMR.040716
|
| [92] |
Jamal SA, Hamilton CJ (2012) Nitric oxide donors for the treatment of osteoporosis. Curr Osteoporos Rep 10: 86-92. doi: 10.1007/s11914-011-0087-7
|
| [93] |
Lazzarato L, Rolando B, Lolli ML, et al. (2005) Synthesis of NO-donor bisphosphonates and their in-vitro action on bone resorption. J Med Chem 48: 1322-1329. doi: 10.1021/jm040830d
|
| [94] |
Kalyanaraman H, Ramdani G, Joshua J, et al. (2017) A Novel, Direct NO Donor Regulates Osteoblast and Osteoclast Functions and Increases Bone Mass in Ovariectomized Mice. J Bone Miner Res 32: 46-59. doi: 10.1002/jbmr.2909
|
| [95] |
Wimalawansa SJ, Chapa MT, Yallampalli C, et al. (1997) Prevention of corticosteroid-induced bone loss with nitric oxide donor nitroglycerin in male rats. Bone 21: 275-280. doi: 10.1016/S8756-3282(97)00125-7
|
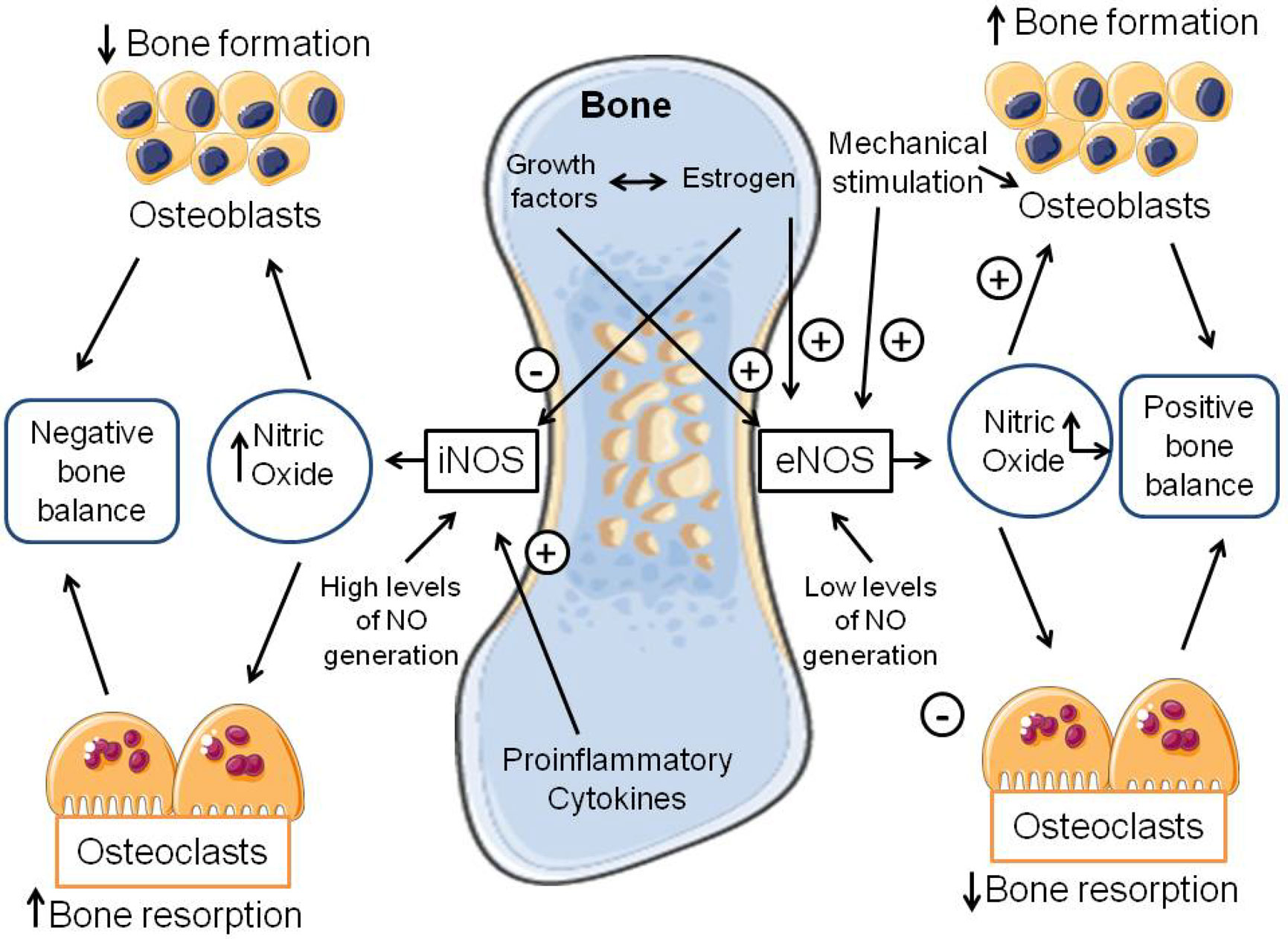
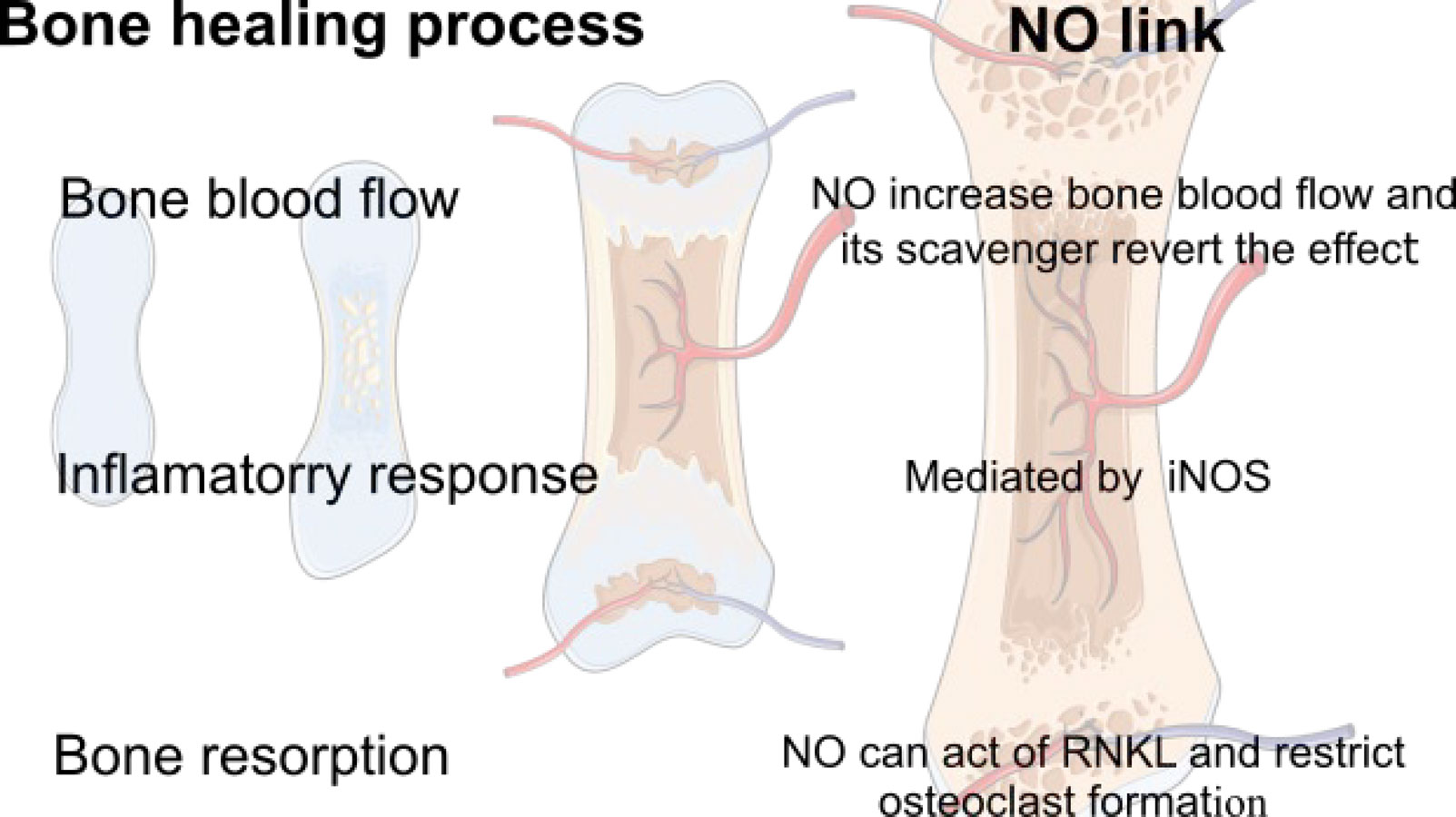
Figures(2) / Tables(1)

Mônica H. M. Nascimento, Milena T. Pelegrino, Joana C. Pieretti, Amedea B. Seabra. How can nitric oxide help osteogenesis?[J]. AIMS Molecular Science, 2020, 7(1): 29-48. doi: 10.3934/molsci.2020003







 DownLoad:
DownLoad: