Citation: Zhaoping Qin, Patrick Robichaud, Taihao Quan. Oxidative stress and CCN1 protein in human skin connective tissue aging[J]. AIMS Molecular Science, 2016, 3(2): 269-279. doi: 10.3934/molsci.2016.2.269

| [1] | Hynes RO, Naba A (2012) Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol 4: a004903. |
| [2] |
Naba A, Clauser KR, Ding H, et al. (2016) The extracellular matrix: Tools and insights for the "omics" era. Matrix Biol 49: 10-24. doi: 10.1016/j.matbio.2015.06.003

|
| [3] | Uitto J (1986) Connective tissue biochemistry of the aging dermis. Age-related alterations in collagen and elastin. Dermatol Clin 4: 433-446. |
| [4] |
Harman D (1981) The aging process. Proc Natl Acad Sci U S A 78: 7124-7128. doi: 10.1073/pnas.78.11.7124
|
| [5] |
Droge W (2002) Free radicals in the physiological control of cell function. Physiol Rev 82: 47-95. doi: 10.1152/physrev.00018.2001
|
| [6] |
Cadenas E, Davies KJ (2000) Mitochondrial free radical generation, oxidative stress, and aging. Free Radic Biol Med 29: 222-230. doi: 10.1016/S0891-5849(00)00317-8
|
| [7] | Fisher GJ, Kang S, Varani J, et al. (2002) Mechanisms of photoaging and chronological skin aging. Arch Dermatol 138: 1462-1470. |
| [8] |
Fisher GJ, Quan T, Purohit T, et al. (2009) Collagen fragmentation promotes oxidative stress and elevates matrix metalloproteinase-1 in fibroblasts in aged human skin. Am J Pathol 174: 101-114. doi: 10.2353/ajpath.2009.080599
|
| [9] | Thomas DR, Burkemper NM (2013) Aging skin and wound healing. Clin Geriatr Med 29: xi-xx. |
| [10] | Worley CA (2006) Aging skin and wound healing. Dermatol Nurs 18: 265-266. |
| [11] | Kudravi SA, Reed MJ (2000) Aging, cancer, and wound healing. In Vivo 14: 83-92. |
| [12] |
Bissell MJ, Kenny PA, Radisky DC (2005) Microenvironmental regulators of tissue structure and function also regulate tumor induction and progression: the role of extracellular matrix and its degrading enzymes. Cold Spring Harb Symp Quant Biol 70: 343-356. doi: 10.1101/sqb.2005.70.013
|
| [13] | Bissell MJ, Hines WC (2011) Why don't we get more cancer? A proposed role of the microenvironment in restraining cancer progression. Nat Med 17: 320-329. |
| [14] |
Fisher GJ, Wang ZQ, Datta SC, et al. (1997) Pathophysiology of premature skin aging induced by ultraviolet light. N Engl J Med 337: 1419-1428. doi: 10.1056/NEJM199711133372003
|
| [15] | Fisher GJ, Varani J, Voorhees JJ (2008) Looking older: fibroblast collapse and therapeutic implications. Arch Dermatol 144: 666-672. |
| [16] |
Yaar M, Gilchrest BA (2001) Skin aging: postulated mechanisms and consequent changes in structure and function. Clin Geriatr Med 17: 617-630. doi: 10.1016/S0749-0690(05)70089-6
|
| [17] |
Quan T, Fisher GJ (2015) Role of Age-Associated Alterations of the Dermal Extracellular Matrix Microenvironment in Human Skin Aging: A Mini-Review. Gerontology 61: 427-434. doi: 10.1159/000371708
|
| [18] |
Yaar M, Eller MS, Gilchrest BA (2002) Fifty years of skin aging. J Investig Dermatol Symp Proc 7: 51-58. doi: 10.1046/j.1523-1747.2002.19636.x
|
| [19] |
Chung JH (2003) Photoaging in Asians. Photodermatol Photoimmunol Photomed 19: 109-121. doi: 10.1034/j.1600-0781.2003.00027.x
|
| [20] |
Wlaschek M, Tantcheva-Poor I, Naderi L, et al. (2001) Solar UV irradiation and dermal photoaging. J Photochem Photobiol B 63: 41-51. doi: 10.1016/S1011-1344(01)00201-9
|
| [21] | Uitto J, Bernstein EF (1998) Molecular mechanisms of cutaneous aging: connective tissue alterations in the dermis. J Investig Dermatol Symp Proc 3: 41-44. |
| [22] |
Lavker RM (1979) Structural alterations in exposed and unexposed aged skin. J Invest Dermatol 73: 59-66. doi: 10.1111/1523-1747.ep12532763
|
| [23] |
Jacob MP (2003) Extracellular matrix remodeling and matrix metalloproteinases in the vascular wall during aging and in pathological conditions. Biomed Pharmacother 57: 195-202. doi: 10.1016/S0753-3322(03)00065-9
|
| [24] |
Cheresh DA, Stupack DG (2008) Regulation of angiogenesis: apoptotic cues from the ECM. Oncogene 27: 6285-6298. doi: 10.1038/onc.2008.304
|
| [25] | Eaglstein WH (1989) Wound healing and aging. Clin Geriatr Med 5: 183-188. |
| [26] |
Valencia IC, Falabella A, Kirsner RS, et al. (2001) Chronic venous insufficiency and venous leg ulceration. J Am Acad Dermatol 44: 401-421; quiz 422-404. doi: 10.1067/mjd.2001.111633
|
| [27] |
Achyut BR, Bader DA, Robles AI, et al. (2013) Inflammation-mediated genetic and epigenetic alterations drive cancer development in the neighboring epithelium upon stromal abrogation of TGF-beta signaling. PLoS Genet 9: e1003251. doi: 10.1371/journal.pgen.1003251
|
| [28] |
Bhowmick NA, Chytil A, Plieth D, et al. (2004) TGF-beta signaling in fibroblasts modulates the oncogenic potential of adjacent epithelia. Science 303: 848-851. doi: 10.1126/science.1090922
|
| [29] |
Mittapalli VR, Madl J, Loffek S, et al. (2016) Injury-Driven Stiffening of the Dermis Expedites Skin Carcinoma Progression. Cancer Res 76: 940-951. doi: 10.1158/0008-5472.CAN-15-1348
|
| [30] |
Fisher GJ, Datta SC, Talwar HS, et al. (1996) Molecular basis of sun-induced premature skin ageing and retinoid antagonism. Nature 379: 335-339. doi: 10.1038/379335a0
|
| [31] |
Yaar M, Gilchrest BA (2007) Photoageing: mechanism, prevention and therapy. Br J Dermatol 157: 874-887. doi: 10.1111/j.1365-2133.2007.08108.x
|
| [32] |
Jung K, Seifert M, Herrling T, et al. (2008) UV-generated free radicals (FR) in skin: their prevention by sunscreens and their induction by self-tanning agents. Spectrochim Acta A Mol Biomol Spectrosc 69: 1423-1428. doi: 10.1016/j.saa.2007.09.029
|
| [33] |
Quan C, Cho MK, Perry D, et al. (2015) Age-associated reduction of cell spreading induces mitochondrial DNA common deletion by oxidative stress in human skin dermal fibroblasts: implication for human skin connective tissue aging. J Biomed Sci 22: 62. doi: 10.1186/s12929-015-0167-6
|
| [34] | He T, Quan T, Shao Y, et al. (2014) Oxidative exposure impairs TGF-β pathway via reduction of type II receptor and SMAD3 in human skin fibroblasts. Age: 1-16. |
| [35] |
Qin Z, Robichaud P, He T, et al. (2014) Oxidant exposure induces cysteine-rich protein 61 (CCN1) via c-Jun/AP-1 to reduce collagen expression in human dermal fibroblasts. PLoS One 9: e115402. doi: 10.1371/journal.pone.0115402
|
| [36] |
Lau LF, Lam SC (1999) The CCN family of angiogenic regulators: the integrin connection. Exp Cell Res 248: 44-57. doi: 10.1006/excr.1999.4456
|
| [37] |
Planque N, Perbal B (2003) A structural approach to the role of CCN (CYR61/CTGF/NOV) proteins in tumourigenesis. Cancer Cell Int 3: 15. doi: 10.1186/1475-2867-3-15
|
| [38] |
Perbal B (2004) CCN proteins: multifunctional signalling regulators. Lancet 363: 62-64. doi: 10.1016/S0140-6736(03)15172-0
|
| [39] |
Leask A, Abraham DJ (2006) All in the CCN family: essential matricellular signaling modulators emerge from the bunker. J Cell Sci 119: 4803-4810. doi: 10.1242/jcs.03270
|
| [40] |
Perbal B, Brigstock DR, Lau LF (2003) Report on the second international workshop on the CCN family of genes. Mol Pathol 56: 80-85. doi: 10.1136/mp.56.2.80
|
| [41] |
Brigstock DR, Goldschmeding R, Katsube KI, et al. (2003) Proposal for a unified CCN nomenclature. Mol Pathol 56: 127-128. doi: 10.1136/mp.56.2.127
|
| [42] |
Chen CC, Lau LF (2009) Functions and mechanisms of action of CCN matricellular proteins. Int J Biochem Cell Biol 41: 771-783. doi: 10.1016/j.biocel.2008.07.025
|
| [43] |
Perumal S, Antipova O, Orgel JP (2008) Collagen fibril architecture, domain organization, and triple-helical conformation govern its proteolysis. Proc Natl Acad Sci U S A 105: 2824-2829. doi: 10.1073/pnas.0710588105
|
| [44] |
Kireeva ML, Mo FE, Yang GP, et al. (1996) Cyr61, a product of a growth factor-inducible immediate-early gene, promotes cell proliferation, migration, and adhesion. Mol Cell Biol 16: 1326-1334. doi: 10.1128/MCB.16.4.1326
|
| [45] |
Chen CC, Chen N, Lau LF (2001) The angiogenic factors Cyr61 and connective tissue growth factor induce adhesive signaling in primary human skin fibroblasts. J Biol Chem 276: 10443-10452. doi: 10.1074/jbc.M008087200
|
| [46] |
Quan T, Qin Z, Shao Y, et al. (2011) Retinoids suppress cysteine-rich protein 61 (CCN1), a negative regulator of collagen homeostasis, in skin equivalent cultures and aged human skin in vivo. Exp Dermatol 20: 572-576. doi: 10.1111/j.1600-0625.2011.01278.x
|
| [47] |
Mo FE, Muntean AG, Chen CC, et al. (2002) CYR61 (CCN1) is essential for placental development and vascular integrity. Mol Cell Biol 22: 8709-8720. doi: 10.1128/MCB.22.24.8709-8720.2002
|
| [48] |
Holbourn KP, Acharya KR, Perbal B (2008) The CCN family of proteins: structure-function relationships. Trends Biochem Sci 33: 461-473. doi: 10.1016/j.tibs.2008.07.006
|
| [49] |
Grzeszkiewicz TM, Kirschling DJ, Chen N, et al. (2001) CYR61 stimulates human skin fibroblast migration through Integrin alpha vbeta 5 and enhances mitogenesis through integrin alpha vbeta 3, independent of its carboxyl-terminal domain. J Biol Chem 276: 21943-21950. doi: 10.1074/jbc.M100978200
|
| [50] |
Kireeva ML, Lam SC, Lau LF (1998) Adhesion of human umbilical vein endothelial cells to the immediate-early gene product Cyr61 is mediated through integrin alphavbeta3. J Biol Chem 273: 3090-3096. doi: 10.1074/jbc.273.5.3090
|
| [51] |
Quan T, He T, Shao Y, et al. (2006) Elevated cysteine-rich 61 mediates aberrant collagen homeostasis in chronologically aged and photoaged human skin. Am J Pathol 169: 482-490. doi: 10.2353/ajpath.2006.060128
|
| [52] |
Quan T, He T, Kang S, et al. (2002) Connective tissue growth factor: expression in human skin in vivo and inhibition by ultraviolet irradiation. J Invest Dermatol 118: 402-408. doi: 10.1046/j.0022-202x.2001.01678.x
|
| [53] |
Quan T, Qin Z, Robichaud P, et al. (2011) CCN1 contributes to skin connective tissue aging by inducing age-associated secretory phenotype in human skin dermal fibroblasts. J Cell Commun Signal 5: 201-207. doi: 10.1007/s12079-011-0144-0
|
| [54] |
Quan T, Qin Z, Xu Y, et al. (2010) Ultraviolet irradiation induces CYR61/CCN1, a mediator of collagen homeostasis, through activation of transcription factor AP-1 in human skin fibroblasts. J Invest Dermatol 130: 1697-1706. doi: 10.1038/jid.2010.29
|
| [55] |
Qin Z, Fisher GJ, Quan T (2013) Cysteine-rich protein 61 (CCN1) domain-specific stimulation of matrix metalloproteinase-1 expression through αVβ3 integrin in human skin fibroblasts. J Biol Chem 288: 12386-12394. doi: 10.1074/jbc.M112.424358
|
| [56] | Quan T, Qin Z, Voorhees JJ, et al. (2012) Cysteine-rich protein 61 (CCN1) mediates replicative senescence-associated aberrant collagen homeostasis in human skin fibroblasts. J Cell Biochem. |
| [57] |
Chen CC, Mo FE, Lau LF (2001) The angiogenic factor Cyr61 activates a genetic program for wound healing in human skin fibroblasts. J Biol Chem 276: 47329-47337. doi: 10.1074/jbc.M107666200
|
| [58] |
Quan T, Shin S, Qin Z, et al. (2009) Expression of CCN family of genes in human skin in vivo and alterations by solar-simulated ultraviolet irradiation. J Cell Commun Signal 3: 19-23. doi: 10.1007/s12079-009-0044-8
|
| [59] |
Qin Z, Okubo T, Voorhees JJ, et al. (2014) Elevated cysteine-rich protein 61 (CCN1) promotes skin aging via upregulation of IL-1β in chronically sun-exposed human skin. Age 36: 353-364. doi: 10.1007/s11357-013-9565-4
|
| [60] | Katsube K, Sakamoto K, Tamamura Y, et al. (2009) Role of CCN, a vertebrate specific gene family, in development. Dev Growth Differ 51: 55-67. |
| [61] |
Jun JI, Lau LF (2010) The matricellular protein CCN1 induces fibroblast senescence and restricts fibrosis in cutaneous wound healing. Nat Cell Biol 12: 676-685. doi: 10.1038/ncb2070
|
| [62] |
Franceschi C, Capri M, Monti D, et al. (2007) Inflammaging and anti-inflammaging: a systemic perspective on aging and longevity emerged from studies in humans. Mech Ageing Dev 128: 92-105. doi: 10.1016/j.mad.2006.11.016
|
| [63] |
Bauge C, Legendre F, Leclercq S, et al. (2007) Interleukin-1beta impairment of transforming growth factor beta1 signaling by down-regulation of transforming growth factor beta receptor type II and up-regulation of Smad7 in human articular chondrocytes. Arthritis Rheum 56: 3020-3032. doi: 10.1002/art.22840
|
| [64] |
Nave H, Fryk S, Pabst R (2000) Scholars in the German Anatomical Institute: longitudinal study concerning the number, gender, personnel positions and the academic education. Ann Anat 182: 489-491. doi: 10.1016/S0940-9602(00)80065-5
|
| [65] |
Giancotti FG, Ruoslahti E (1999) Integrin signaling. Science 285: 1028-1032. doi: 10.1126/science.285.5430.1028
|
| [66] |
Mocsai A, Abram CL, Jakus Z, et al. (2006) Integrin signaling in neutrophils and macrophages uses adaptors containing immunoreceptor tyrosine-based activation motifs. Nat Immunol 7: 1326-1333. doi: 10.1038/ni1407
|
| [67] |
Berrier AL, Yamada KM (2007) Cell-matrix adhesion. J Cell Physiol 213: 565-573. doi: 10.1002/jcp.21237
|
| [68] |
Chakraborti S, Mandal M, Das S, et al. (2003) Regulation of matrix metalloproteinases: an overview. Mol Cell Biochem 253: 269-285. doi: 10.1023/A:1026028303196
|
| [69] | Gutman A, Wasylyk B (1990) The collagenase gene promoter contains a TPA and oncogene-responsive unit encompassing the PEA3 and AP-1 binding sites. EMBO J 9: 2241-2246. |
| [70] |
Benbow U, Brinckerhoff CE (1997) The AP-1 site and MMP gene regulation: what is all the fuss about? Matrix Biol 15: 519-526. doi: 10.1016/S0945-053X(97)90026-3
|
| [71] | Foletta VC, Segal DH, Cohen DR (1998) Transcriptional regulation in the immune system: all roads lead to AP-1. J Leukoc Biol 63: 139-152. |
| [72] | Fisher G, Datta S, Wang Z, et al. (2000) c-Jun dependent inhibition of cutaneous procollagen transcription following ultraviolet irradiation is reversed by all-trans retinoid acid. J Clin Invest 106: 661-668. |
| [73] |
Verrecchia F, Pessah M, Atfi A, et al. (2000) Tumor necrosis factor-a inhibits transforming growth factor-ß/Smad signaling in human dermal fibroblasts via AP-1 activation. J Biol Chem 275: 30226-30231. doi: 10.1074/jbc.M005310200
|
| [74] | Dennler S, Prunier C, Ferrand N, et al. (2000) c-Jun inhibits transforming growth factor beta-mediated transcription by repressing Smad3 transcriptional activity. J Biol Chem 275: 28858-28865. |
| [75] |
Qin Z, Fisher GJ, Quan T (2013) Cysteine-rich protein 61 (CCN1) domain-specific stimulation of matrix metalloproteinase-1 expression through alphaVbeta3 integrin in human skin fibroblasts. J Biol Chem 288: 12386-12394. doi: 10.1074/jbc.M112.424358
|
| [76] |
Qin Z, Okubo T, Voorhees JJ, et al. (2014) Elevated cysteine-rich protein 61 (CCN1) promotes skin aging via upregulation of IL-1beta in chronically sun-exposed human skin. Age 36: 353-364. doi: 10.1007/s11357-013-9565-4
|
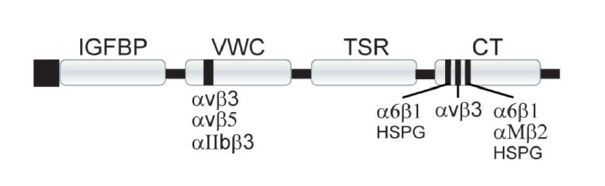
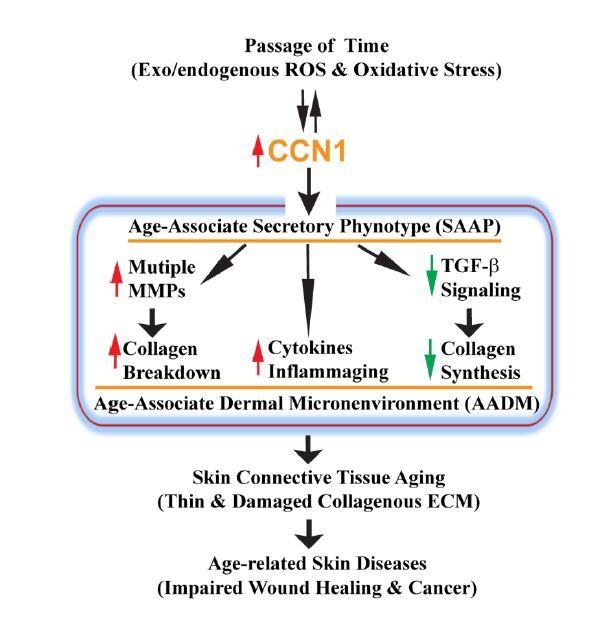
Figures(2)

Zhaoping Qin, Patrick Robichaud, Taihao Quan. Oxidative stress and CCN1 protein in human skin connective tissue aging[J]. AIMS Molecular Science, 2016, 3(2): 269-279. doi: 10.3934/molsci.2016.2.269







 DownLoad:
DownLoad: