Citation: Brion Woroch, Alex Konkel, Brian D. Gonsalves. Activation of stimulus-specific processing regions at retrieval tracks the strength of relational memory[J]. AIMS Neuroscience, 2019, 6(4): 250-265. doi: 10.3934/Neuroscience.2019.4.250

| [1] | Cohen NJ, Eichenbaum H (1993) Memory, amnesia, and the hippocampal system. Cambridge, MA, US: The MIT Press. |
| [2] | Eichenbaum H, Cohen NJ (2001) From conditioning to conscious recollection: Memory systems of the brain. New York: Oxford University Press. |
| [3] |
Rissman J, Wagner AD (2012) Distributed representations in memory: insights from functional brain imaging. Annu Rev Psychol 63: 101–128. doi: 10.1146/annurev-psych-120710-100344

|
| [4] |
Danker JF, Anderson JR (2010) The ghosts of brain states past: remembering reactivates the brain regions engaged during encoding. Psychol Bull 136: 87–102. doi: 10.1037/a0017937
|
| [5] | Rugg MD, Vilberg KL (2014) Brain Networks Underlying Episodic Memory Retrieval. Curr Opin Neurobilogy 23: 255–260. |
| [6] | Morcom AM (2014) Re-engaging with the past: recapitulation of encoding operations during episodic retrieval. Front Hum Neurosci 8: 351. |
| [7] |
Wheeler ME, Petersen SE, Buckner RL (2000) Memory's echo: Vivid remembering reactivates sensory-specific cortex. Proc Natl Acad Sci 97: 11125–11129. doi: 10.1073/pnas.97.20.11125
|
| [8] |
Johnson JD, McDuff SGR, Rugg MD, et al. (2009) Recollection, familiarity, and cortical reinstatement: a multivoxel pattern analysis. Neuron 63: 697–708. doi: 10.1016/j.neuron.2009.08.011
|
| [9] | Thakral PP, Wang TH, Rugg MD (2017)Decoding the content of recollection within the core recollection network and beyond. Cortex 91: 101–113. |
| [10] |
Johnson JD, Rugg MD (2007) Recollection and the reinstatement of encoding-related cortical activity. Cereb Cortex 17: 2507–2515. doi: 10.1093/cercor/bhl156
|
| [11] |
Alvarez P, Squire LR (1994) Memory consolidation and the medial temporal lobe: A simple network model. Proc Natl Acad Sci U S A 91: 7041–7045. doi: 10.1073/pnas.91.15.7041
|
| [12] |
McClelland JL, McNaughton BL, O'Reilly RC (1995) Why there are complementary learning systems in the hippocampus and neocortex: insights from the successes and failures of connectionist models of learning and memory. Psychol Rev 102: 419–457. doi: 10.1037/0033-295X.102.3.419
|
| [13] |
Moscovitch M, Rosenbaum RS, Gilboa A, et al. (2005) Functional neuroanatomy of remote episodic, semantic and spatial memory: A unified account based on multiple trace theory. J Anat 207: 35–66. doi: 10.1111/j.1469-7580.2005.00421.x
|
| [14] |
Norman KA, O'Reilly RC (2003) Modeling hippocampal and neocortical contributions to recognition memory: A complementary-learning-systems approach. Psychol Rev 110: 611–646. doi: 10.1037/0033-295X.110.4.611
|
| [15] |
Rosler F, Heil M, Hennighausen E (1995) Distinct Cortical Activation Patterns during Long-Term Memory Retrieval of Verbal , Spatial , and Color Information. J Cogn Neurosci 7: 51–65. doi: 10.1162/jocn.1995.7.1.51
|
| [16] |
Khader P, Burke M, Bien S, et al. (2005) Content-specific activation during associative long-term memory retrieval. Neuroimage 27: 805–816. doi: 10.1016/j.neuroimage.2005.05.006
|
| [17] |
Nyberg L, Habib R, Herlitz A (2000) Brain activation during episodic memory retrieval: sex differences. Acta Psychol (Amst) 105: 181–194. doi: 10.1016/S0001-6918(00)00060-3
|
| [18] |
Vaidya CJ, Zhao M, Desmond JE, et al. (2002) Evidence for cortical encoding specificity in episodic memory: memory-induced re-activation of picture processing areas. Neuropsychologia 40: 2136–2143. doi: 10.1016/S0028-3932(02)00053-2
|
| [19] |
Wheeler ME, Buckner RL (2003) Functional dissociation among components of remembering: control, perceived oldness, and content. J Neurosci 23: 3869–3880. doi: 10.1523/JNEUROSCI.23-09-03869.2003
|
| [20] |
Wheeler ME, Buckner RL (2004) Functional-anatomic correlates of remembering and knowing. Neuroimage 21: 1337–1349. doi: 10.1016/j.neuroimage.2003.11.001
|
| [21] |
Woodruff CC, Johnson JD, Uncapher MR, et al. (2005) Content-specificity of the neural correlates of recollection. Neuropsychologia 43: 1022–1032. doi: 10.1016/j.neuropsychologia.2004.10.013
|
| [22] |
Liang JC, Preston AR (2017) Medial temporal lobe reinstatement of content-specific details predicts source memory. Cortex 91: 67–78. doi: 10.1016/j.cortex.2016.09.011
|
| [23] | Leiker EK, Johnson JD (2015) Pattern reactivation co-varies with activity in the core recollection network during source memory. Neuropsychologia 75. |
| [24] |
Maratos EJ, Dolan RJ, Morris JS, et al. (2001) Neural activity associated with episodic memory for emotional context. Neuropsychologia 39: 910–920. doi: 10.1016/S0028-3932(01)00025-2
|
| [25] |
Smith AP, Henson RN, Dolan RJ, et al. (2004) fMRI correlates of the episodic retrieval of emotional contexts. Neuroimage 22: 868–878. doi: 10.1016/j.neuroimage.2004.01.049
|
| [26] |
Kahn I, Davachi L, Wagner AD (2004) Functional-neuroanatomic correlates of recollection: implications for models of recognition memory. J Neurosci 24: 4172–4180. doi: 10.1523/JNEUROSCI.0624-04.2004
|
| [27] |
Voss JL, Galvan A, Gonsalves BD (2011) Neuropsychologia Cortical regions recruited for complex active-learning strategies and action planning exhibit rapid reactivation during memory retrieval. Neuropsychologia 49: 3956–3966. doi: 10.1016/j.neuropsychologia.2011.10.012
|
| [28] |
Dodson CS, Holland PW, Shimamura AP (1998) On the recollection of specific-and partial-source information. J Exp Psychol Learn Mem Cogn 24: 1121–1136. doi: 10.1037/0278-7393.24.5.1121
|
| [29] |
Hicks JL, Marsh RL, Ritschel L (2002) The role of recollection and partial information in source monitoring. J Exp Psychol Learn Mem Cogn 28: 503–508. doi: 10.1037/0278-7393.28.3.503
|
| [30] |
Simons JS, Dodson CS, Bell D, et al. (2004) Specific- and partial-source memory: effects of aging. Psychol Aging 19: 689–694. doi: 10.1037/0882-7974.19.4.689
|
| [31] |
Slotnick SD, Dodson CS (2005) Support for a continuous (single-process) model of recognition memory and source memory. Mem Cognit 33: 151–170. doi: 10.3758/BF03195305
|
| [32] |
Mickes L, Wais P, Wixted J (2009) Recollection is a continuous process implications for dual-process theories of recognition memory. Psychol Sci 20: 509–515. doi: 10.1111/j.1467-9280.2009.02324.x
|
| [33] |
Slotnick SD (2010) Remember' source memory ROCs indicate recollection is a continuous process. Memory 18: 27–39. doi: 10.1080/09658210903390061
|
| [34] |
Vilberg KL, Rugg MD (2008) Memory retrieval and the parietal cortex: A review of evidence from a dual-process perspective. Neuropsychologia 46: 1787–1799. doi: 10.1016/j.neuropsychologia.2008.01.004
|
| [35] |
Vilberg KL, Rugg MD (2009) Left parietal cortex is modulated by amount of recollected verbal information. Neuroreport 20: 1295–1299. doi: 10.1097/WNR.0b013e3283306798
|
| [36] |
Vilberg KL, Rugg MD (2007) Dissociation of the neural correlates of recognition memory according to familiarity, recollection, and amount of recollected information. Neuropsychologia 45: 2216–2225. doi: 10.1016/j.neuropsychologia.2007.02.027
|
| [37] |
Shimamura AP (2011) Episodic retrieval and the cortical binding of relational activity. Cogn Affect Behav Neurosci 11: 277–291. doi: 10.3758/s13415-011-0031-4
|
| [38] | Thakral PP, Wang TH, Rugg MD (2015) Cortical reinstatement and the confidence and accuracy of source memory. Neuroimage 109. |
| [39] |
Mack ML, Preston AR (2016) Decisions about the past are guided by reinstatement of specific memories in the hippocampus and perirhinal cortex. Neuroimage 127: 144–157. doi: 10.1016/j.neuroimage.2015.12.015
|
| [40] |
Ishai A (2008) Let's face it: it's a cortical network. Neuroimage 40: 415–419. doi: 10.1016/j.neuroimage.2007.10.040
|
| [41] |
Epstein R, Kanwisher N (1998) A cortical representation of the local visual environment. Nature 392: 598–601. doi: 10.1038/33402
|
| [42] |
O'Craven KM, Kanwisher N (2000) Mental imagery of faces and places activates corresponding stiimulus-specific brain regions. J Cogn Neurosci 12: 1013–1023. doi: 10.1162/08989290051137549
|
| [43] |
Althoff RR, Cohen NJ (1999) Eye-movement-based memory effect: A reprocessing effect in face perception. J Exp Psychol Learn Mem Cogn 25: 997–1010. doi: 10.1037/0278-7393.25.4.997
|
| [44] |
Walther DB, Caddigan E, Fei-Fei L, et al. (2009) Natural scene categories revealed in distributed patterns of activity in the human brain. J Neurosci 29: 10573–10581. doi: 10.1523/JNEUROSCI.0559-09.2009
|
| [45] | Brett M, Anton J, Valabregue R, et al. (2002) Region of interest analysis using an SPM toolbox [abstract] Presented at the 8th International Conference on Functional Mapping of the Human Brain, June 2-6, 2002, Sendai, Japan. Available on CD-ROM in NeuroImage, Vol 16. |
| [46] |
Warriner AB, Kuperman V, Brysbaert M (2013) Norms of valence, arousal, and dominance for 13,915 English lemmas. Behav Res Methods 45: 1191–1207. doi: 10.3758/s13428-012-0314-x
|
| [47] |
Diana RA, Yonelinas AP, Ranganath C (2007) Imaging recollection and familiarity in the medial temporal lobe: A three-component model. Trends Cogn Sci 11: 379–386. doi: 10.1016/j.tics.2007.08.001
|
| [48] |
Gonsalves BD, Kahn I, Curran T, et al. (2005) Memory strength and repetition suppression: multimodal imaging of medial temporal cortical contributions to recognition. Neuron 47: 751–761. doi: 10.1016/j.neuron.2005.07.013
|
| [49] |
Wais PE (2008) FMRI signals associated with memory strength in the medial temporal lobes: A meta-analysis. Neuropsychologia 46: 3185–3196. doi: 10.1016/j.neuropsychologia.2008.08.025
|

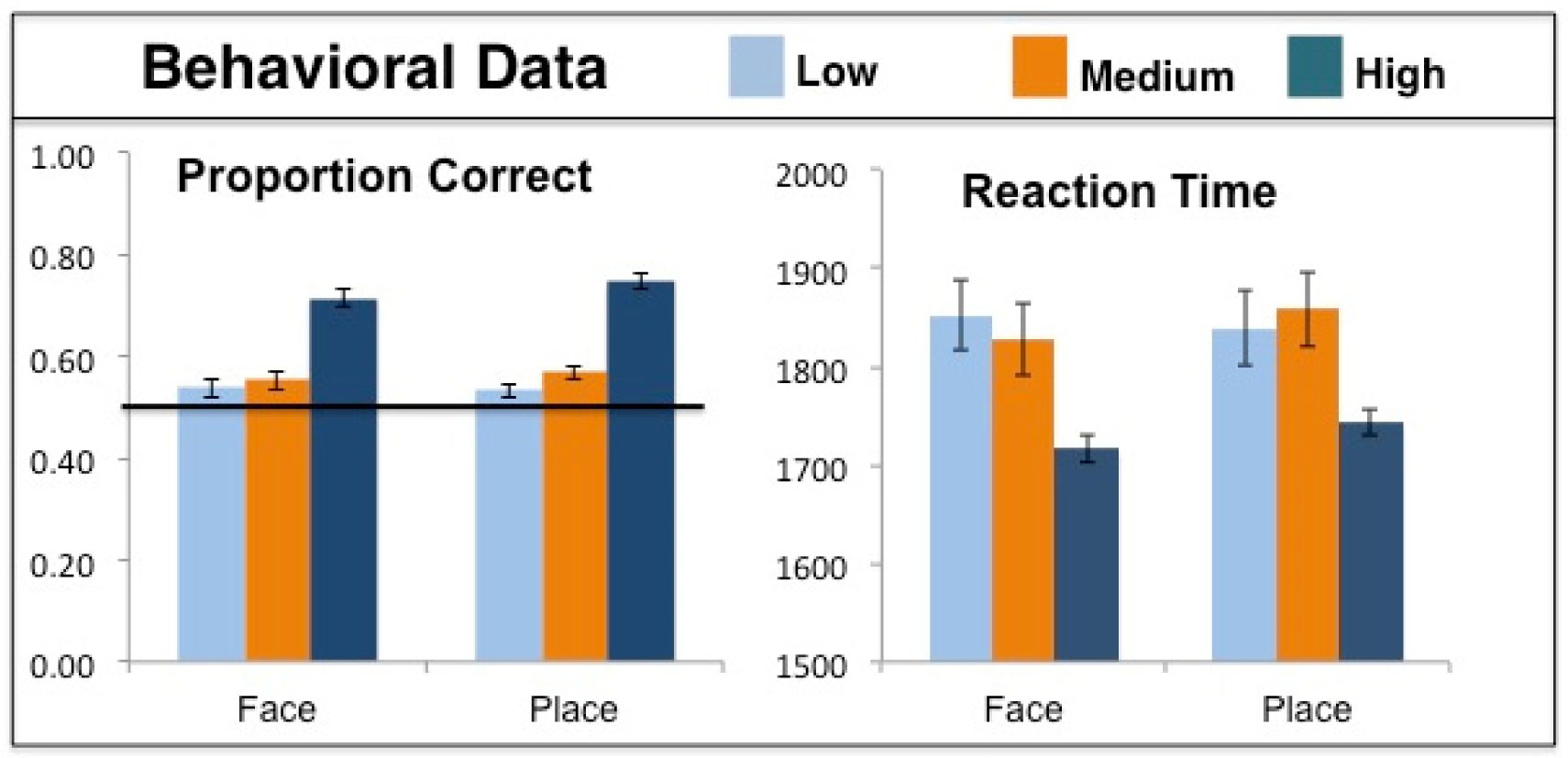
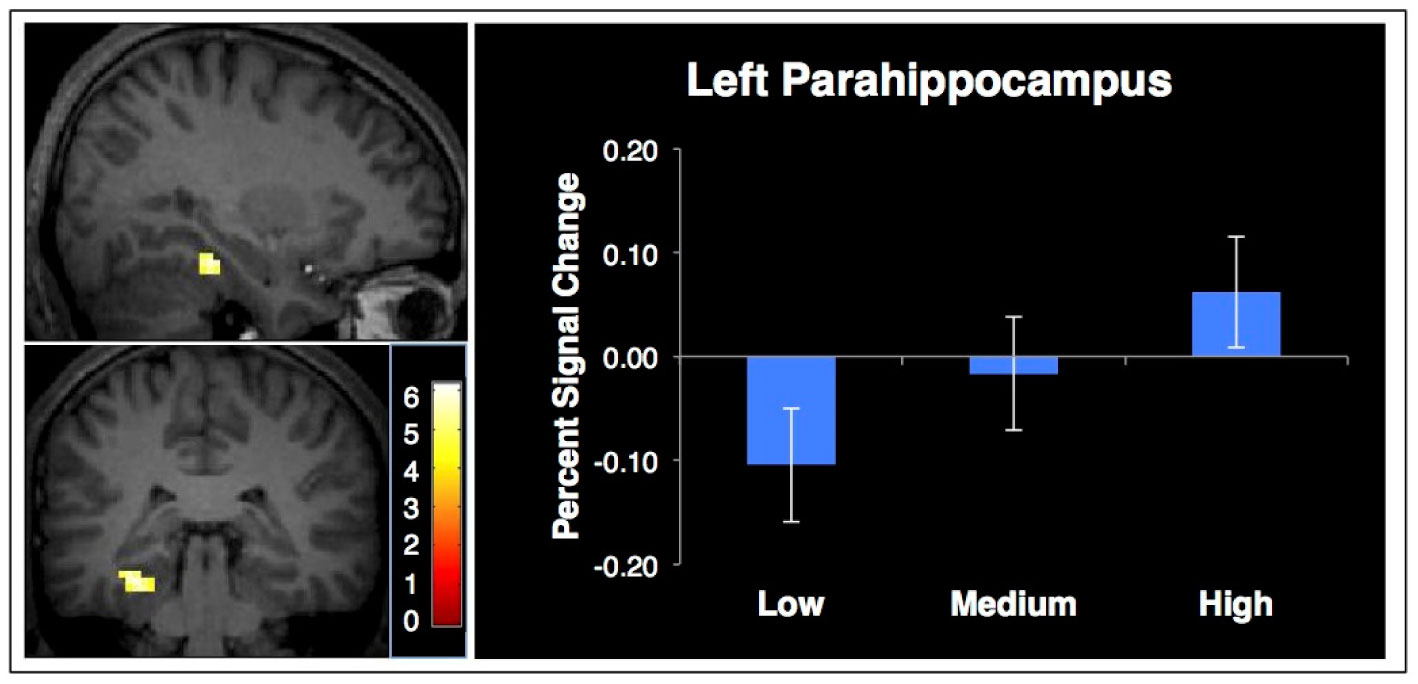
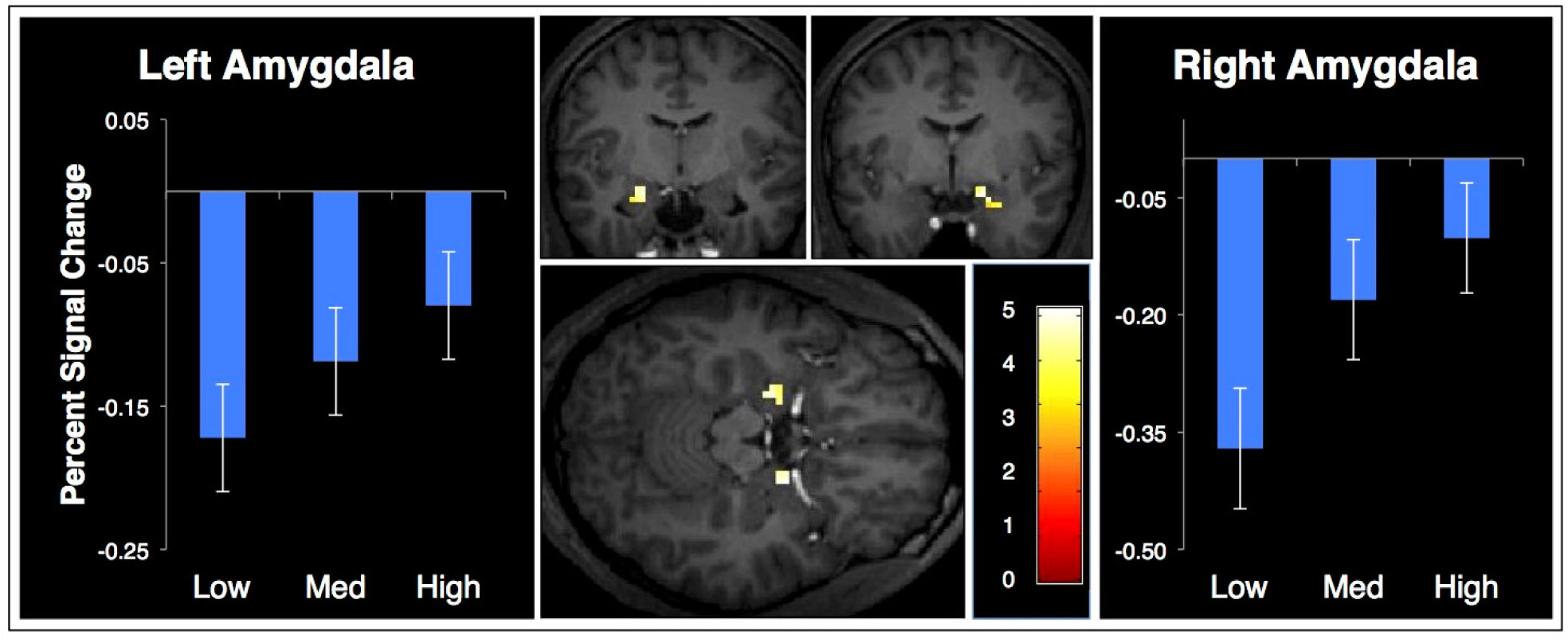
Figures(4) / Tables(1)

Brion Woroch, Alex Konkel, Brian D. Gonsalves. Activation of stimulus-specific processing regions at retrieval tracks the strength of relational memory[J]. AIMS Neuroscience, 2019, 6(4): 250-265. doi: 10.3934/Neuroscience.2019.4.250







 DownLoad:
DownLoad: