Citation: Amory H. Danek, Virginia L. Flanagin. Cognitive conflict and restructuring: The neural basis of two core components of insight[J]. AIMS Neuroscience, 2019, 6(2): 60-84. doi: 10.3934/Neuroscience.2019.2.60

| [1] | Danek AH (2018) Magic tricks, sudden restructuring and the Aha! experience: A new model of non-monotonic problem solving, In: Vallée-Tourangeau F (ed), Insight: On the origins of new ideas, London: Routledge, 51–78. |
| [2] | Danek AH, Wiley J, Öllinger M (2016) Solving classical insight problems without Aha! experience: 9 Dot, 8 Coin, and Matchstick Arithmetic Problems. J Probl Solving 9: 47–57. |
| [3] | Bowden EM (1997) The effect of reportable and unreportable hints on anagram solution and the Aha! experience. Conscious Cogn 6: 545–573. |
| [4] |
Cushen PJ, Wiley J (2012) Cues to solution, restructuring patterns, and reports of insight in creative problem solving. Conscious Cogn 21: 1166–1175. doi: 10.1016/j.concog.2012.03.013

|
| [5] |
Stroop JR (1935) Studies of interference in serial verbal reactions. J Exp Psychol 18: 643–662. doi: 10.1037/h0054651
|
| [6] |
Luo J, Knoblich G (2007) Studying insight problem solving with neuroscientific methods. Methods 42: 77–86. doi: 10.1016/j.ymeth.2006.12.005
|
| [7] | Ohlsson S (1984) Restructuring revisited: II. An information processing theory of restructuring and insight. Scand J Psychol 25: 117–129. |
| [8] | Smith SM (1995) Getting into and out of mental ruts: A theory of fixation, incubation, and insight, In: Sternberg RJ, Davidson JE (eds), The nature of insight, Cambridge, MA: MIT Press, 229–251. |
| [9] | Wertheimer M (1925) Über Schlussprozesse im produktiven Denken. In: Wertheimer M (ed), Drei Abhandlungen zur Gestalttheorie, Erlangen: Verlag der Philosophischen Akademie, 164–184. |
| [10] |
Knoblich G, Ohlsson S, Haider H, et al. (1999) Constraint relaxation and chunk decomposition in insight problem solving. J Exp Psychol Learn Mem Cogn 25: 1534–1555. doi: 10.1037/0278-7393.25.6.1534
|
| [11] | Ohlsson S (1992) Information-processing explanations of insight and related phenomena, In: Keane MT, Gilhooly KJ (eds), Advances in the psychology of thinking, London: Harvester-Wheatsheaf, 1–44. |
| [12] | Ohlsson S (2011) Deep learning: How the mind overrides experience, New York: Cambridge University Press. |
| [13] | Sandkühler S, Bhattacharya J (2008) Deconstructing insight: EEG correlates of insightful problem solving. PLoS One 3: 1–12. |
| [14] |
Smith RW, Kounios J (1996) Sudden insight: All-or-none processing revealed by speed-accuracy decomposition. J Exp Psychol Learn Mem Cogn 22: 1443–1462. doi: 10.1037/0278-7393.22.6.1443
|
| [15] |
Metcalfe J (1986) Feeling of knowing in memory and problem solving. J Exp Psychol Learn Mem Cogn 12: 288–294. doi: 10.1037/0278-7393.12.2.288
|
| [16] |
Metcalfe J, Wiebe D (1987) Intuition in insight and noninsight problem solving. Mem Cognit 15: 238–246. doi: 10.3758/BF03197722
|
| [17] |
Tik M, Sladky R, Luft CDB, et al. (2018) Ultra-high-field fMRI insights on insight: Neural correlates of the Aha!-moment. Hum Brain Mapp 39: 3241–3252. doi: 10.1002/hbm.24073
|
| [18] | Jung-Beeman M, Bowden EM, Haberman J, et al. (2004) Neural activity when people solve verbal problems with insight. PLoS Biol 2: 500–510. |
| [19] |
Starchenko MG, Bekhtereva NP, Pakhomov SV, et al. (2003) Study of the brain organization of creative thinking. Hum Physiol 29: 652–653. doi: 10.1023/A:1025836521833
|
| [20] |
Bechtereva NP, Korotkov AD, Pakhomov SV, et al. (2004) PET study of brain maintenance of verbal creative activity. Int J Psychophysiol 53: 11–20. doi: 10.1016/j.ijpsycho.2004.01.001
|
| [21] |
Luo J, Niki K, Phillips S (2004) Neural correlates of the Aha! reaction. NeuroReport 15: 2013–2017. doi: 10.1097/00001756-200409150-00004
|
| [22] |
Dietrich A, Kanso R (2010) A review of EEG, ERP, and neuroimaging studies of creativity and insight. Psychol Bull 136: 822–848. doi: 10.1037/a0019749
|
| [23] |
Bowden EM, Jung-Beeman M (2003) Aha! Insight experience correlates with solution activation in the right hemisphere. Psychon Bull Rev 10: 730–737. doi: 10.3758/BF03196539
|
| [24] |
Kounios J, Beeman M (2014) The cognitive neuroscience of insight. Annu Rev Psychol 65: 71–93. doi: 10.1146/annurev-psych-010213-115154
|
| [25] |
Sprugnoli G, Rossi S, Emmendorfer A, et al. (2017) Neural correlates of Eureka moment. Intelligence 62: 99–118. doi: 10.1016/j.intell.2017.03.004
|
| [26] | Luo J, Niki K, Phillips S (2004) The function of the anterior cingulate cortex (ACC) in the insightful solving of puzzles: The ACC is activated less when the structure of the puzzle is known. J Psychol Chin Soc 5: 195–213. |
| [27] |
Zhao Q, Zhou Z, Xu H, et al. (2013) Dynamic neural network of insight: a functional magnetic resonance imaging study on solving chinese 'chengyu' riddles. PloS One 8: e59351. doi: 10.1371/journal.pone.0059351
|
| [28] |
Aziz-Zadeh L, Kaplan JT, Iacoboni M (2009) "Aha!": The neural correlates of verbal insight solutions. Hum Brain Mapp 30: 908–916. doi: 10.1002/hbm.20554
|
| [29] | Danek AH, Fraps T, von Müller A, et al. (2014) Working wonders? Investigating insight with magic tricks. Cognition 130: 174–185. |
| [30] | Danek AH, Öllinger M, Fraps T, et al. (2015) An fMRI investigation of expectation violation in magic tricks. Front Psychol 6: 48. |
| [31] |
Danek AH, Fraps T, von Müller A, et al. (2013) Aha! experiences leave a mark: facilitated recall of insight solutions. Psychol Res 77: 659–669. doi: 10.1007/s00426-012-0454-8
|
| [32] | Ekroll V, Sayim B, Wagemans J (2013) Against better knowledge: The magical force of amodal volume completion. Iperception 4: 511–515. |
| [33] |
Ekroll V, Sayim B, Wagemans J (2017) The other side of magic: the psychology of perceiving hidden things. Perspect Psychol Sci 12: 91–106. doi: 10.1177/1745691616654676
|
| [34] |
Ekroll V, Sayim B, Van der Hallen R, et al. (2016) Illusory visual completion of an object's invisible backside can make your finger feel shorter. Curr Biol 26: 1029–1033. doi: 10.1016/j.cub.2016.02.001
|
| [35] | Pétervári J, Danek AH (under review) Problem solving of magic tricks: Guiding to and through an impasse with solution cues. |
| [36] | Danek AH, Wiley J (2017) What about false insights? Deconstructing the Aha! experience along its multiple dimensions for correct and incorrect solutions separately. Front Psychol 7: 2077. |
| [37] |
Koo TK, Li MY (2016) A guideline of selecting and reporting intraclass correlation coefficients for reliability research. J Chiropr Med 15: 155–163. doi: 10.1016/j.jcm.2016.02.012
|
| [38] |
Nichols TE (2012) Multiple testing corrections, nonparametric methods, and random field theory. NeuroImage 62: 811–815. doi: 10.1016/j.neuroimage.2012.04.014
|
| [39] |
Woo C-W, Krishnan A, Wager TD (2014) Cluster-extent based thresholding in fMRI analyses: Pitfalls and recommendations. NeuroImage 91: 412–419. doi: 10.1016/j.neuroimage.2013.12.058
|
| [40] |
Eickhoff SB, Stephan KE, Mohlberg H, et al. (2005) A new SPM toolbox for combining probabilistic cytoarchitectonic maps and functional imaging data. NeuroImage 25: 1325–1335. doi: 10.1016/j.neuroimage.2004.12.034
|
| [41] |
Auzias G, Coulon O, Brovelli A (2016) MarsAtlas: A cortical parcellation atlas for functional mapping. Hum Brain Mapp 37: 1573–1592. doi: 10.1002/hbm.23121
|
| [42] |
Carlén M (2017) What constitutes the prefrontal cortex? Science 358: 478–482. doi: 10.1126/science.aan8868
|
| [43] |
Nee DE, Brown JW, Askren MK, et al. (2013) A meta-analysis of executive components of working memory. Cereb Cortex 23: 264–282. doi: 10.1093/cercor/bhs007
|
| [44] |
Lobel E, Kahane P, Leonards U, et al. (2001) Localization of human frontal eye fields: anatomical and functional findings of functional magnetic resonance imaging and intracerebral electrical stimulation. J Neurosurg 95: 804–815. doi: 10.3171/jns.2001.95.5.0804
|
| [45] |
Caspers S, Eickhoff SB, Geyer S, et al. (2008) The human inferior parietal lobule in stereotaxic space. Brain Struct Funct 212: 481–495. doi: 10.1007/s00429-008-0195-z
|
| [46] |
Thomas C, Didierjean A (2016) Magicians fix your mind: How unlikely solutions block obvious ones. Cognition 154: 169–173. doi: 10.1016/j.cognition.2016.06.002
|
| [47] |
Thomas C, Didierjean A, Kuhn G (2018) It is magic! How impossible solutions prevent the discovery of obvious ones? Q J Exp Psychol 71: 2481–2487. doi: 10.1177/1747021817743439
|
| [48] | Wright RD, Lawrence MW (2008) Orienting of attention, Oxford: Oxford University Press. |
| [49] |
Moore T, Fallah M (2004) Microstimulation of the frontal eye field and its effects on covert spatial attention. J Neurophysiol 91: 152–162. doi: 10.1152/jn.00741.2002
|
| [50] |
Wang H, Callaghan E, Gooding-Williams G, et al. (2016) Rhythm makes the world go round: An MEG-TMS study on the role of right TPJ theta oscillations in embodied perspective taking. Cortex 75: 68–81. doi: 10.1016/j.cortex.2015.11.011
|
| [51] | Vernet M, Quentin R, Chanes L, et al. (2014) Frontal eye field, where art thou? Anatomy, function, and non-invasive manipulation of frontal regions involved in eye movements and associated cognitive operations. Front Integr Neurosci 8: 66. |
| [52] |
Greicius MD, Krasnow B, Reiss AL, et al. (2003) Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proc Natl Acad Sci 100: 253–258. doi: 10.1073/pnas.0135058100
|
| [53] |
Kizilirmak JM, Schott BH, Thuerich H, et al. (2019) Learning of novel semantic relationships via sudden comprehension is associated with a hippocampus-independent network. Conscious Cogn 69: 113–132. doi: 10.1016/j.concog.2019.01.005
|
| [54] |
Gilmore AW, Nelson SM, McDermott KB (2015) A parietal memory network revealed by multiple MRI methods. Trends Cogn Sci 19: 534–543. doi: 10.1016/j.tics.2015.07.004
|
| [55] |
Zacks JM, Speer NK, Swallow KM, et al. (2007) Event perception: A mind-brain perspective. Psychol Bull 133: 273–293. doi: 10.1037/0033-2909.133.2.273
|
| [56] |
Albright TD (2012) On the perception of probable things: neural substrates of associative memory, imagery, and perception. Neuron 74: 227–245. doi: 10.1016/j.neuron.2012.04.001
|
| [57] |
Ochsner KN, Hughes B, Robertson ER, et al. (2009) Neural systems supporting the control of affective and cognitive conflicts. J Cogn Neurosci 21: 1841–1854. doi: 10.1162/jocn.2009.21129
|
| [58] |
Botvinick MM (2007) Conflict monitoring and decision making: Reconciling two perspectives on anterior cingulate function. Cogn Affect Behav Neurosci 7: 356–366. doi: 10.3758/CABN.7.4.356
|
| [59] | Kizilirmak JM, Thuerich H, Folta-Schoofs K, et al. (2016) Neural correlates of learning from induced insight: a case for reward-based episodic encoding. Front Psychol 7: 1693. |
| [60] |
Dandan T, Haixue Z, Wenfu L, et al. (2013) Brain activity in using heuristic prototype to solve insightful problems. Behav Brain Res 253: 139–144. doi: 10.1016/j.bbr.2013.07.017
|
| [61] |
Seghier ML (2013) The Angular Gyrus: Multiple Functions and Multiple Subdivisions. The Neuroscientist 19: 43–61. doi: 10.1177/1073858412440596
|
| [62] | Binder JR, Desai RH, Graves WW, et al. (2009) Where is the semantic system? A critical review and meta-analysis of 120 functional neuroimaging studies. Cereb Cortex 19: 2767–2796. |
| [63] |
Vandenberghe R, Price C, Wise R, et al. (1996) Functional anatomy of a common semantic system for words and pictures. Nature 383: 254–6. doi: 10.1038/383254a0
|
| [64] |
Ye Z, Zhou X (2009) Conflict control during sentence comprehension: fMRI evidence. Neuroimage 48: 280–90. doi: 10.1016/j.neuroimage.2009.06.032
|
| [65] |
Spratling MW (2016) Predictive coding as a model of cognition. Cogn Process 17: 279–305. doi: 10.1007/s10339-016-0765-6
|
| [66] | Benn Y, Webb TL, Chang BPI, et al. (2014) The neural basis of monitoring goal progress. Front Hum Neurosci 8: 688. |
| [67] | Craig AD (2009) How do you feel-now? The anterior insula and human awareness. Nat Rev Neurosci 10: 59–70. |
| [68] |
Tian F, Tu S, Qiu J, et al. (2011) Neural correlates of mental preparation for successful insight problem solving. Behav Brain Res 216: 626–630. doi: 10.1016/j.bbr.2010.09.005
|
| [69] | Boccia M, Piccardi L, Palermo L, et al. (2015) Where do bright ideas occur in our brain? Meta-analytic evidence from neuroimaging studies of domain-specific creativity. Front Psychol 6: 1195. |
| [70] |
Sridharan D, Levitin DJ, Menon V (2008) A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks. Proc Natl Acad Sci 105: 12569. doi: 10.1073/pnas.0800005105
|
| [71] |
Beaty RE, Benedek M, Barry Kaufman S, et al. (2015) Default and executive network coupling supports creative idea production. Sci Rep 5: 10964. doi: 10.1038/srep10964
|
| [72] | Menon V (2015) Salience Network. Brain Mapp Encycl Ref 2: 597–611. |
| [73] |
Parris BA, Kuhn G, Mizon GA, et al. (2009) Imaging the impossible: An fMRI study of impossible causal relationships in magic tricks. NeuroImage 45: 1033–1039. doi: 10.1016/j.neuroimage.2008.12.036
|
| [74] |
Grahn JA, Parkinson JA, Owen AM (2008) The cognitive functions of the caudate nucleus. Prog Neurobiol 86: 141–155. doi: 10.1016/j.pneurobio.2008.09.004
|
| [75] |
Schneider M, Leuchs L, Czisch M, et al. (2018) Disentangling reward anticipation with simultaneous pupillometry/fMRI. Neuroimage 178: 11–22. doi: 10.1016/j.neuroimage.2018.04.078
|


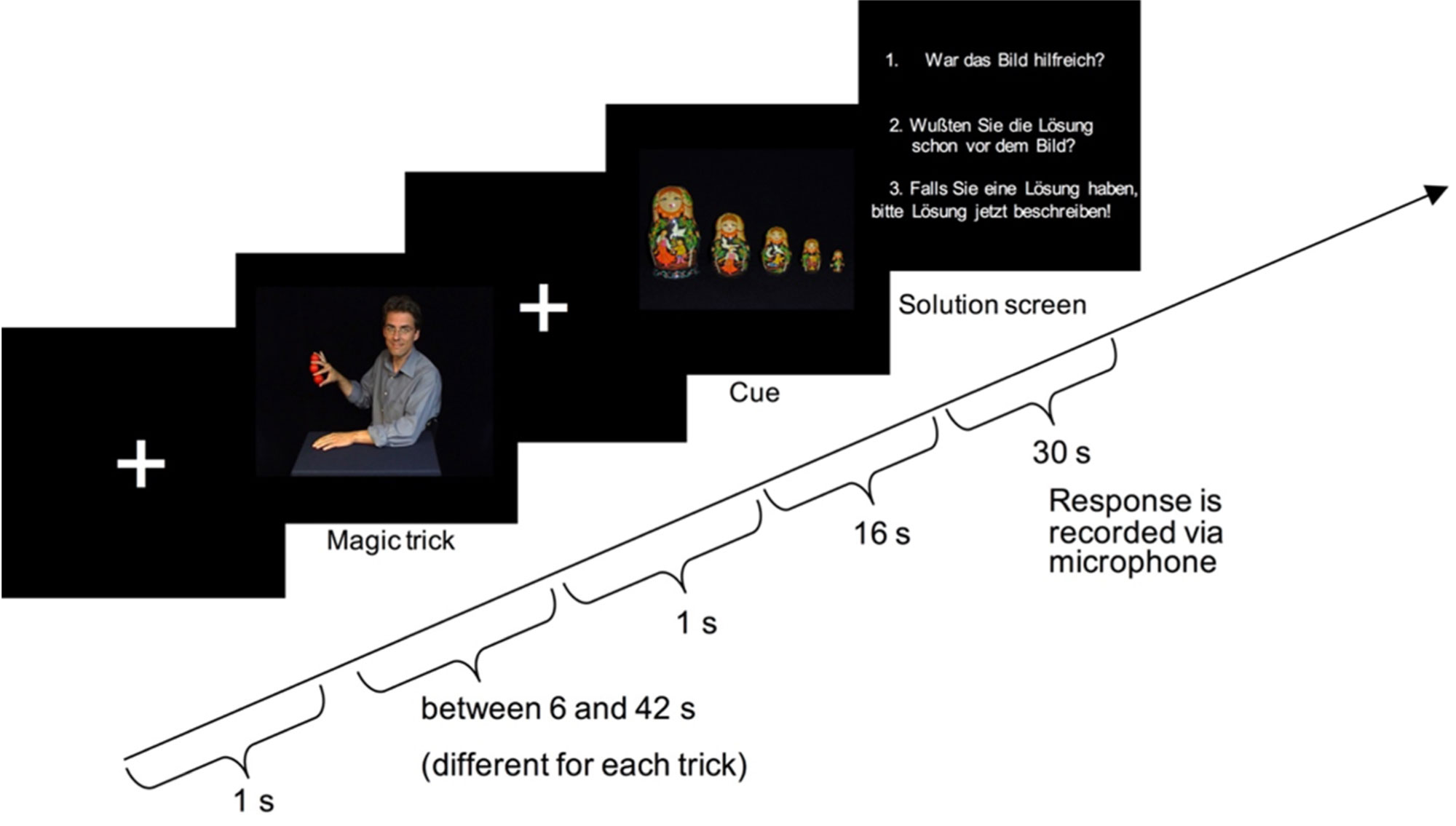
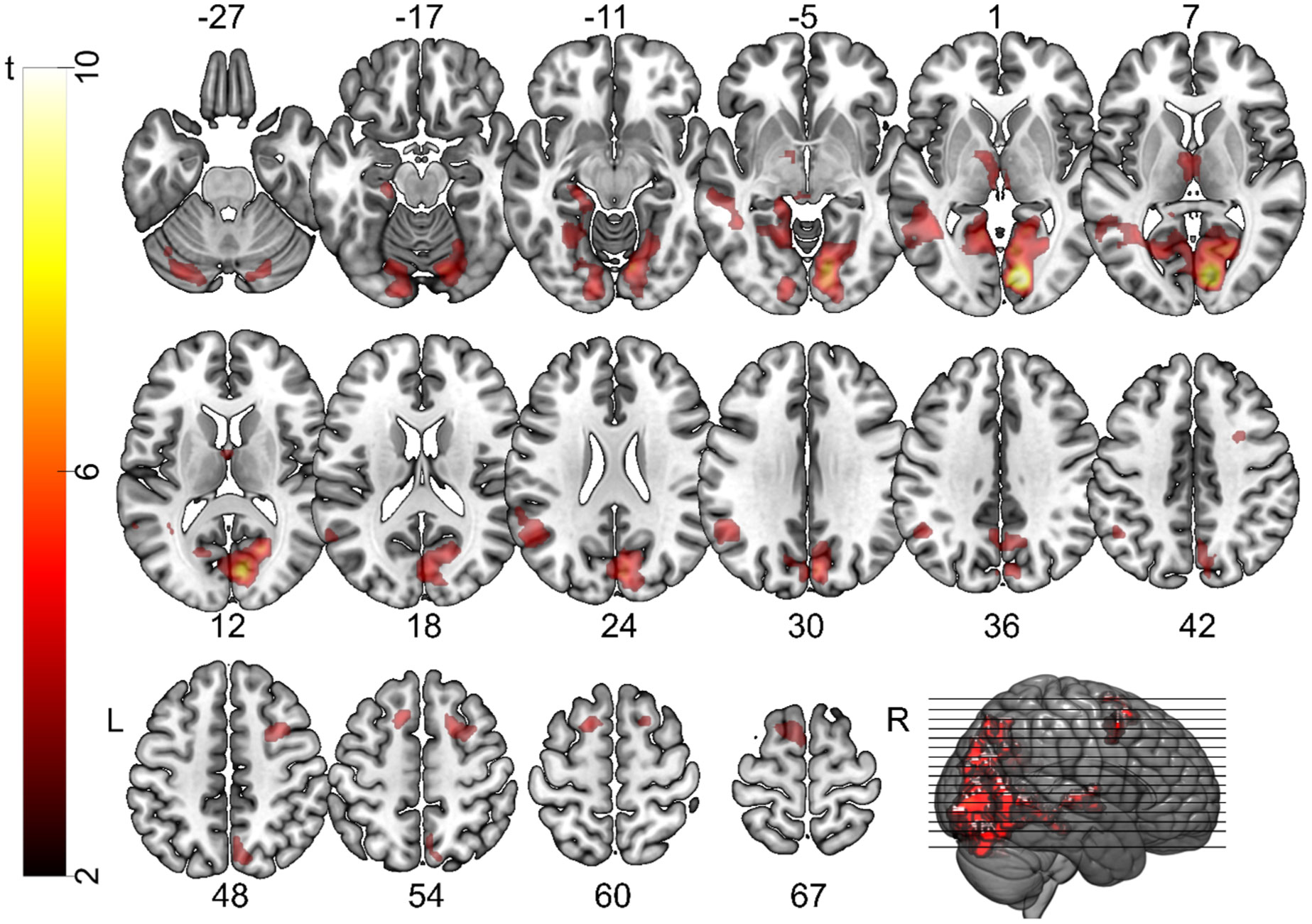
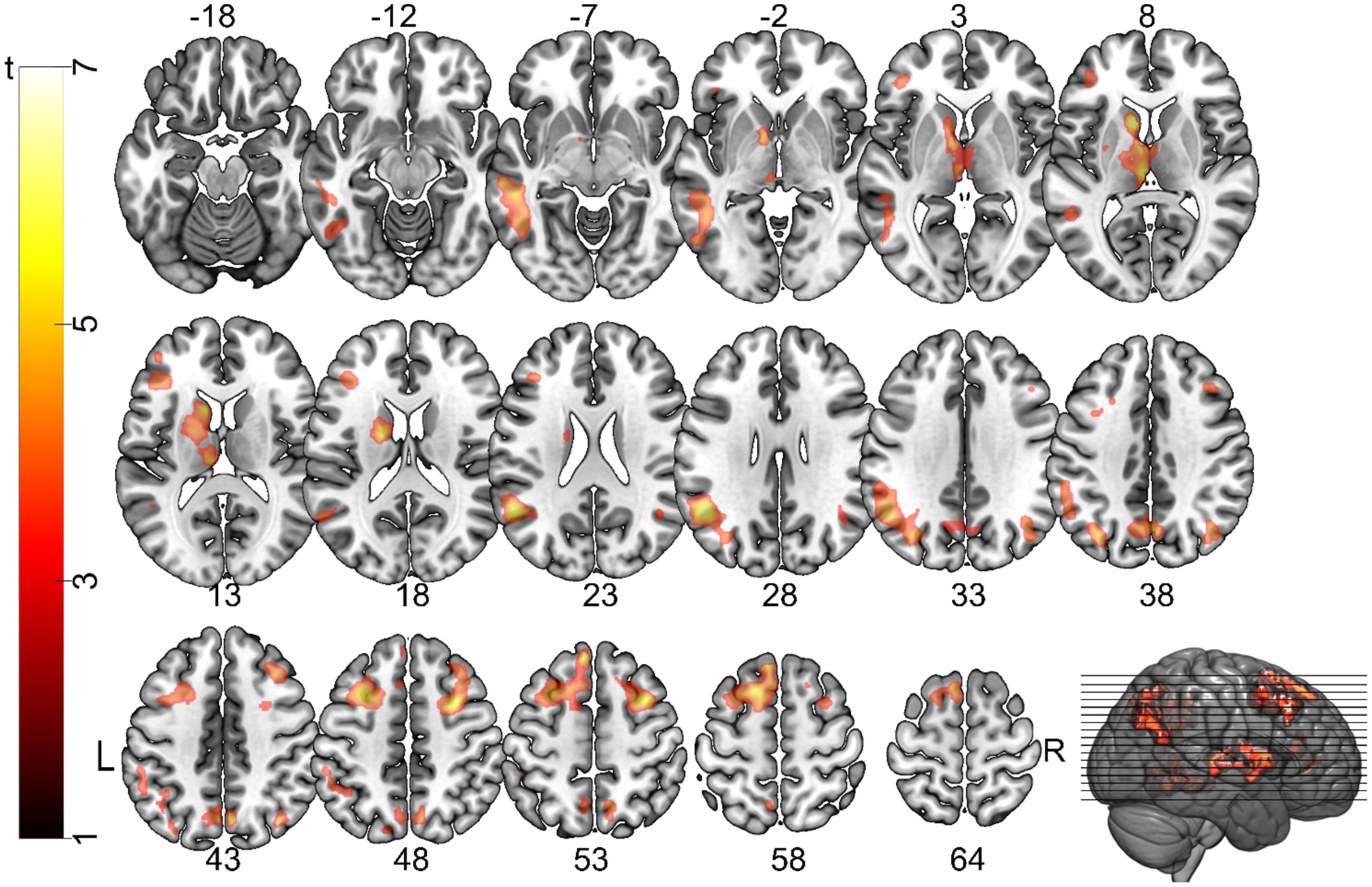
Figures(5) / Tables(2)

Amory H. Danek, Virginia L. Flanagin. Cognitive conflict and restructuring: The neural basis of two core components of insight[J]. AIMS Neuroscience, 2019, 6(2): 60-84. doi: 10.3934/Neuroscience.2019.2.60







 DownLoad:
DownLoad: