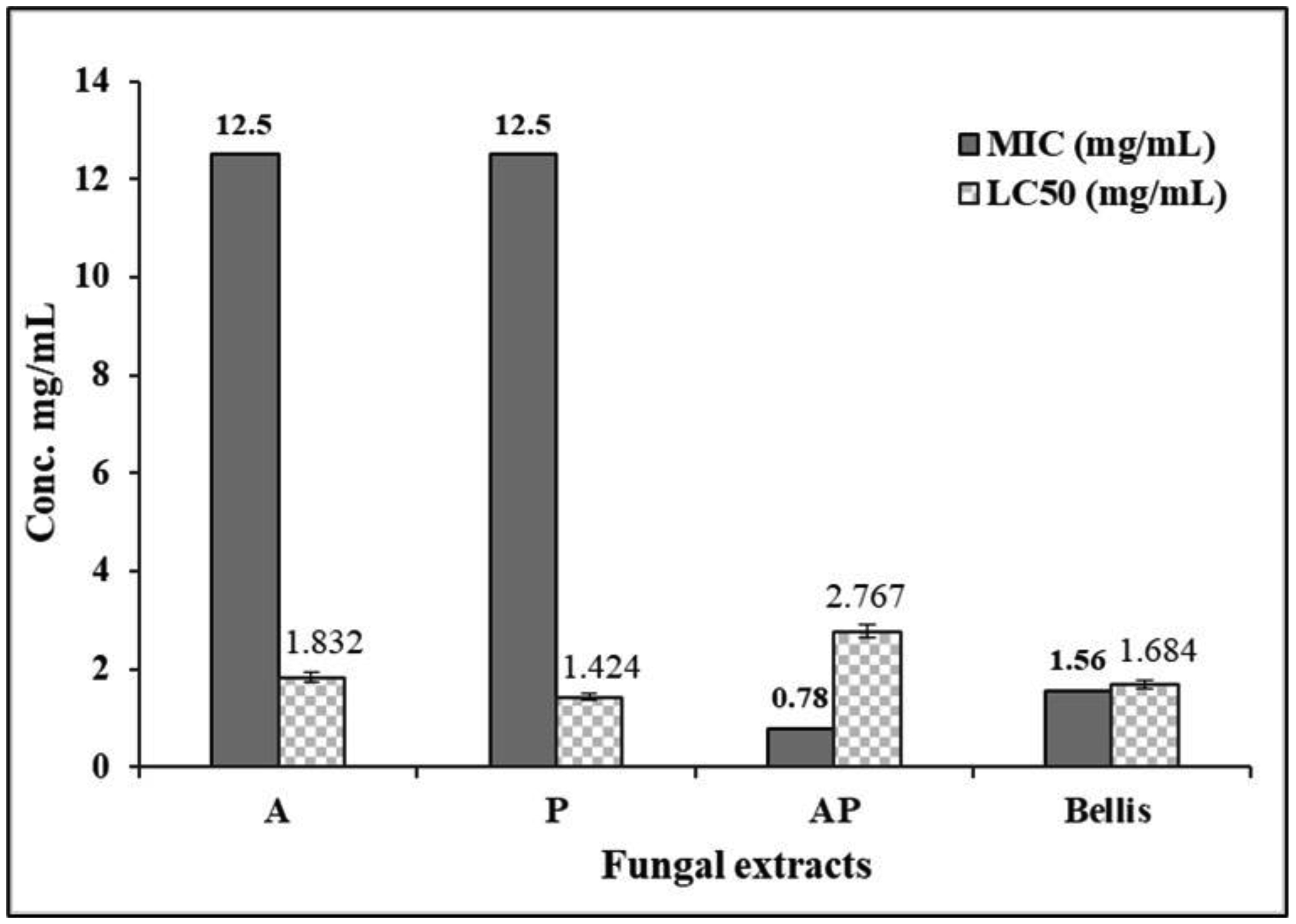
Fungal co-culture is a method that allows the detection of interactions between fungi, enabling the examination of bioactive novel metabolites induction that may not be produced in monocultures. Worldwide, Fusarium basal rot is a primary limitation to onion yield, being caused by different Fusarium species. Current research directions encourage biological control of plant diseases as a replacement for routine chemical treatments. The current study aimed to investigate the co-culturing technique for mining new sources of bioagents that could be used as fungicides. Aspergillus ochraceus AUMC15539 was co-cultured with Penicillium chrysogenum AUMC15504, and their ethyl acetate extract was tested in vitro and in a greenhouse against Fusarium proliferatum AUMC15541. The results showed that Aspergillus-Penicillium (AP) co-culture extract significantly inhibited the growth of F. proliferatum with an MIC value of 0.78 mg/mL and showed antioxidant efficiency with an IC50 value of 1.31 mg/mL. The brine shrimp toxicity testing showed a LC50 value of 2.77 mg/mL. In addition, the co-culture extract showed the highest phenolic content at 114.71 GAE mg/g, with a 27.82 QE mg/g flavonoid content. Profiling of AP co-culture and its monoculture extracts by HPLC revealed a change in the metabolites profile in AP co-culture. Principal component analysis verified a positive correlation between the obtained HPLC data of A. ochraceus (A), P. chrysogenum (P), and AP extracts. Greenhouse experiments demonstrated that treating infected onion plants with the AP co-culture extract significantly enhanced all growth parameters. Additionally, the co-culture extract treatment resulted in the highest levels of total pigments (3.46 mg/g), carbohydrates (52.10 mg/g dry weight), proteins (131.44 mg/g), phenolics (41.66 GAE mg/g), and flavonoids (9.43 QE mg/g) compared with other treatments. This indicates a promising potential for fungal co-cultures in discovering new bioagents with antifungal properties and growth-promoting capabilities.
Citation: Mohammed M. M. Abdelrahem, Mohamed E. Abouelela, Nageh F. Abo-Dahab, Abdallah M. A. Hassane. Aspergillus-Penicillium co-culture: An investigation of bioagents for controlling Fusarium proliferatum-induced basal rot in onion[J]. AIMS Microbiology, 2024, 10(4): 1024-1051. doi: 10.3934/microbiol.2024044
Fungal co-culture is a method that allows the detection of interactions between fungi, enabling the examination of bioactive novel metabolites induction that may not be produced in monocultures. Worldwide, Fusarium basal rot is a primary limitation to onion yield, being caused by different Fusarium species. Current research directions encourage biological control of plant diseases as a replacement for routine chemical treatments. The current study aimed to investigate the co-culturing technique for mining new sources of bioagents that could be used as fungicides. Aspergillus ochraceus AUMC15539 was co-cultured with Penicillium chrysogenum AUMC15504, and their ethyl acetate extract was tested in vitro and in a greenhouse against Fusarium proliferatum AUMC15541. The results showed that Aspergillus-Penicillium (AP) co-culture extract significantly inhibited the growth of F. proliferatum with an MIC value of 0.78 mg/mL and showed antioxidant efficiency with an IC50 value of 1.31 mg/mL. The brine shrimp toxicity testing showed a LC50 value of 2.77 mg/mL. In addition, the co-culture extract showed the highest phenolic content at 114.71 GAE mg/g, with a 27.82 QE mg/g flavonoid content. Profiling of AP co-culture and its monoculture extracts by HPLC revealed a change in the metabolites profile in AP co-culture. Principal component analysis verified a positive correlation between the obtained HPLC data of A. ochraceus (A), P. chrysogenum (P), and AP extracts. Greenhouse experiments demonstrated that treating infected onion plants with the AP co-culture extract significantly enhanced all growth parameters. Additionally, the co-culture extract treatment resulted in the highest levels of total pigments (3.46 mg/g), carbohydrates (52.10 mg/g dry weight), proteins (131.44 mg/g), phenolics (41.66 GAE mg/g), and flavonoids (9.43 QE mg/g) compared with other treatments. This indicates a promising potential for fungal co-cultures in discovering new bioagents with antifungal properties and growth-promoting capabilities.

| [1] |
Newman DJ, Cragg GM (2012) Natural products as sources of new drugs over the 30 years from 1981 to 2010. J Nat Prod 75: 311-335. https://doi.org/10.1021/np200906s 
|
| [2] |
Akondi KB, Lakshmi VV (2013) Emerging trends in genomic approaches for microbial bioprospecting. Omics 17: 61-70. https://doi.org/10.1089/omi.2012.0082
|
| [3] |
Demain AL (2014) Importance of microbial natural products and the need to revitalize their discovery. J Ind Microbiol Biotechnol 41: 185-201. https://doi.org/10.1007/s10295-013-1325-z
|
| [4] |
Ola ARB, Thomy D, Lai D, et al. (2013) Inducing secondary metabolite production by the endophytic fungus Fusarium tricinctum through coculture with Bacillus subtilis. J Nat Prod 76: 2094-2099. https://doi.org/10.1021/np400589h
|
| [5] | Powthong P, Jantrapanukorn B, Thongmee A, et al. (2013) Screening of antimicrobial activities of the endophytic fungi isolated from Sesbania grandiflora (L.) Pers. J Agr Sci Tech 15: 1513-1522. |
| [6] |
Singh A, Singh DK, Kharwar RN, et al. (2021) Fungal endophytes as efficient sources of plant-derived bioactive compounds and their prospective applications in natural product drug discovery: Insights, avenues, and challenges. Microorganisms 9: 197. https://doi.org/10.3390/microorganisms9010197
|
| [7] | Verma VC, Kharwar RN, Strobel GA (2009) Chemical and functional diversity of natural products from plant associated endophytic fungi. Nat Prod Commun 4: 1511-1532. https://doi.org/10.1177/1934578X0900401114 |
| [8] |
Sharaf MH, Abdelaziz AM, Kalaba MH, et al. (2022) Antimicrobial, antioxidant, cytotoxic activities and phytochemical analysis of fungal endophytes isolated from Ocimum Basilicum. Appl Biochem Biotechnol 194: 1271-1289. https://doi.org/10.1007/s12010-021-03702-w
|
| [9] |
Weyens N, van der Lelie D, Taghavi S, et al. (2009) Exploiting plant–microbe partnerships to improve biomass production and remediation. Trends Biotechnol 27: 591-598. https://doi.org/10.1016/j.tibtech.2009.07.006
|
| [10] |
Devi R, Kaur T, Kour D, et al. (2020) Beneficial fungal communities from different habitats and their roles in plant growth promotion and soil health. Microb Biosyst 5: 21-47. https://doi.org/10.21608/mb.2020.32802.1016
|
| [11] |
Serrano R, González-Menéndez V, Rodríguez L, et al. (2017) Co-culturing of fungal strains against Botrytis cinerea as a model for the induction of chemical diversity and therapeutic agents. Front Microbiol 8: 649. https://doi.org/10.3389/fmicb.2017.00649
|
| [12] |
Nai C, Meyer V (2018) From axenic to mixed cultures: Technological advances accelerating a paradigm shift in microbiology. Trends Microbiol 26: 538-554. https://doi.org/10.1016/j.tim.2017.11.004
|
| [13] |
Ochi K, Hosaka T (2013) New strategies for drug discovery: activation of silent or weakly expressed microbial gene clusters. Appl Microbiol Biotechnol 97: 87-98. https://doi.org/10.1007/s00253-012-4551-9
|
| [14] |
Marmann A, Aly AH, Lin W, et al. (2014) Co-cultivation—a powerful emerging tool for enhancing the chemical diversity of microorganisms. Mar Drugs 12: 1043-1065. https://doi.org/10.3390/md12021043
|
| [15] |
De Roy K, Marzorati M, Van den Abbeele P, et al. (2014) Synthetic microbial ecosystems: An exciting tool to understand and apply microbial communities. Environ Microbiol 16: 1472-1481. https://doi.org/10.1111/1462-2920.12343
|
| [16] |
Moody SC (2014) Microbial co-culture: Harnessing intermicrobial signaling for the production of novel antimicrobials. Future Microbiol 9: 575-578. https://doi.org/10.2217/fmb.14.25
|
| [17] |
Alabouvette C, Olivain C, Migheli Q, et al. (2009) Microbiological control of soil-borne phytopathogenic fungi with special emphasis on wilt-inducing Fusarium oxysporum. New Phytol 184: 529-544. https://doi.org/10.1111/j.1469-8137.2009.03014.x
|
| [18] |
Mohamed H, Hassane A, Atta O, et al. (2021) Deep learning strategies for active secondary metabolites biosynthesis from fungi: Harnessing artificial manipulation and application. Biocatal Agric Biotechnol 38: 102195. https://doi.org/10.1016/j.bcab.2021.102195
|
| [19] |
Kozlovskii AG, Zhelifonova VP, Antipova TV (2013) Fungi of the genus Penicillium as producers of physiologically active compounds. Appl Biochem Microbiol 49: 1-10. https://doi.org/10.1134/S0003683813010092
|
| [20] |
Sun Y, Liu WC, Shi X, et al. (2021) Inducing secondary metabolite production of Aspergillus sydowii through microbial co-culture with Bacillus subtilis. Microb Cell Fact 20: 1-16. https://doi.org/10.1186/s12934-021-01527-0
|
| [21] |
Gebretsadik K, Dechassa N (2018) Response of onion (Allium cepa L.) to nitrogen fertilizer rates and spacing under rain fed condition at Tahtay Koraro, Ethiopia. Sci Rep 8: 9495. https://doi.org/10.1038/s41598-018-27762-x
|
| [22] |
Omar AE, Al-Khalaifah HS, Mohamed WAM, et al. (2020) Effects of phenolic-rich onion (Allium cepa L.) extract on the growth performance, behavior, intestinal histology, amino acid digestibility, antioxidant activity, and the immune status of broiler chickens. Front Vet Sci 7: 582612. https://doi.org/10.3389/fvets.2020.582612
|
| [23] | Yahaya Y, Uauri UAB, Bagudo BU (2010) Study of nutrient content variation in bulb and stalk of onions (Allium Sepa) cultivated in Aliero, Aliero, Kebbi State, Nigeria. Nigerian J Basic Appl Sci 18: 83-89. |
| [24] |
Attaya E, Bardisi A, Osman A, et al. (2024) Effect of nitrogen fertilization levels and plant density on dry weight, yield components and bulb quality of onion plant. Not Bot Horti Agrobot Cluj-Napoca 52: 13294. https://doi.org/10.15835/nbha52113294
|
| [25] |
Cramer CS (2000) Breeding and genetics of Fusarium basal rot resistance in onion. Euphytica 115: 159-166. https://doi.org/10.1023/A:1004071907642
|
| [26] |
Bayraktar H, Türkkan M, Dolar FS (2010) Characterization of Fusarium oxysporum f. sp. cepae from onion in Turkey based on vegetative compatibility and rDNA RFLP analysis. J Phytopathol 158: 691-697. https://doi.org/10.1111/j.1439-0434.2010.01685.x
|
| [27] |
Fan CM, Xiong GR, Qi P, et al. (2008) Potential biofumigation effects of Brassica oleracea var. caulorapa on growth of fungi. J Phytopathol 156: 321-325. https://doi.org/10.1111/j.1439-0434.2007.01343.x
|
| [28] |
Aktar W, Sengupta D, Chowdhury A (2009) Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip Toxicol 2: 1-12. https://doi.org/10.2478/v10102-009-0001-7
|
| [29] | Rongai D, Pulcini P, Pesce B, et al. (2015) Antifungal activity of some botanical extracts on Fusarium oxysporum. Open Life Sci 10: 409-416. https://doi.org/10.1515/biol-2015-0040 |
| [30] |
Oppong-Danquah E, Budnicka P, Blümel M, et al. (2020) Design of fungal co-cultivation based on comparative metabolomics and bioactivity for discovery of marine fungal agrochemicals. Mar Drugs 18: 73. https://doi.org/10.3390/md18020073
|
| [31] |
Abdelrahem MMM, Hassane AMA, Abouelela ME, et al. (2023) Comparative bioactivity and metabolites produced by fungal co-culture system against myco-phytopathogens. J Environ Stud 31: 1-15. https://doi.org/10.21608/jesj.2023.232560.1056
|
| [32] |
Frisvad JC, Filtenborg O (1983) Classification of terverticillate penicillia based on profiles of mycotoxins and other secondary metabolites. Appl Environ Microbiol 46: 1301-1310. https://doi.org/10.1128/aem.46.6.1301-1310.1983
|
| [33] |
Hassan MM, Farid MA, Gaber A (2019) Rapid identification of Trichoderma koningiopsis and Trichoderma longibrachiatum using sequence-characterized amplified region markers. Egypt J Biol Pest Control 29: 13. https://doi.org/10.1186/s41938-019-0113-0
|
| [34] |
Mohamed H, El-Shanawany AR, Shah AM, et al. (2020) Comparative analysis of different isolated oleaginous Mucoromycota fungi for their γ-linolenic acid and carotenoid production. BioMed Res Int 2020: 3621543. https://doi.org/10.1155/2020/3621543
|
| [35] |
Al Mousa AA, Mohamed H, Hassane AMA, et al. (2021) Antimicrobial and cytotoxic potential of an endophytic fungus Alternaria tenuissima AUMC14342 isolated from Artemisia judaica L. growing in Saudi Arabia. J King Saud Univ Sci 33: 101462. https://doi.org/10.1016/j.jksus.2021.101462
|
| [36] |
Park JH, Gyung JC, Kyoung SJ, et al. (2005) Antifungal activity against plant pathogenic fungi of chaetoviridins isolated from Chaetomium globosum. FEMS Microbiol Lett 252: 309-313. https://doi.org/10.1016/j.femsle.2005.09.013
|
| [37] |
Meyer BN, Ferrigni NR, Putnam JE, et al. (1982) Brine shrimp: A convenient general bioassay for active plant constituents. Planta Medica 45: 31-34. https://doi.org/10.1055/s-2007-971236
|
| [38] | Moshi MJ, Van den Beukel CJ, Hamza OJM, et al. (2007) Brine shrimp toxicity evaluation of some Tanzanian plants used traditionally for the treatment of fungal infections. Afr J Trad Complement Altern Med 4: 219-225. https://doi.org/10.4314/ajtcam.v4i2.31211 |
| [39] |
Kupina S, Fields C, Roman MC, et al. (2018) Determination of total phenolic content using the Folin-C assay: Single-laboratory validation, first action 2017.13. J AOAC Int 101: 1466-1472. https://doi.org/10.5740/jaoacint.18-0031
|
| [40] |
Quettier-Deleu C, Gressier B, Vasseur J, et al. (2000) Phenolic compounds and antioxidant activities of buckwheat (Fagopyrum esculentum Moench) hulls and flour. J Ethnopharmacol 72: 35-42. https://doi.org/10.1016/S0378-8741(00)00196-3
|
| [41] |
Brand-Williams W, Cuvelier ME, Berset C (1995) Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci Technol 28: 25-30. https://doi.org/10.1016/S0023-6438(95)80008-5
|
| [42] |
Hassane AMA, Saleh MH, Mohamed EA, et al. (2022) In vitro and in silico antioxidant efficiency of bio-potent secondary metabolites from different taxa of black seeds producing plants and their derived-mycoendophytes. Front Bioeng Biotechnol 10: 930161. https://doi.org/10.3389/fbioe.2022.930161
|
| [43] | Akpotu MO, Eze PM, Abba CC, et al. (2017) Antimicrobial activities of secondary metabolites of endophytic fungi isolated from Catharanthus roseus. J Health Sci 7: 15-22. https://doi.org/10.17532/jhsci.2017.421 |
| [44] | Riaz T, Khan SN, Javaid A (2010) Management of Fusarium corm rot of gladiolus (Gladiolus grandiflorus sect. Blandus cv. Aarti) by using leaves of allelopathic plants. Afr J Biotechnol 9: 4681-4686. |
| [45] | Hassanein NM, Zeid MAA, Youssef KA, et al. (2010) Control of tomato early blight and wilt using aqueous extract of neem leaves. Phytopathol Mediterr 49: 143-151. https://doi.org/10.14601/Phytopathol_Mediterr-3085 |
| [46] | Johansen LE, Curi EA, Bonf JH, et al. (1960) Methods for Studying Soil Microflora Plant Disease Relationship. Minneapolis: Burge's Publishing Co. 77. |
| [47] |
Boughalleb-M'hamdi N, Salem IB, M'hamdi M (2018) Evaluation of the efficiency of Trichoderma, Penicillium, and Aspergillus species as biological control agents against four soil-borne fungi of melon and watermelon. Egypt J Biol Pest Control 28: 1-12. https://doi.org/10.1186/s41938-017-0010-3
|
| [48] |
Ajmal M, Ahmad S, Hussain S (2001) Effect of soil moisture on black scurf disease and yield in potato. Pak J Biol Sci 4: 175-201.
|
| [49] |
Metwally RA, Al-Amri SM (2020) Individual and interactive role of Trichoderma viride and arbuscular mycorrhizal fungi on growth and pigment content of onion plants. Lett Appl Microbiol 70: 79-86. https://doi.org/10.1111/lam.13246
|
| [50] |
Lichtenthaler HK, Wellburn AR (1983) Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem Soc Trans 11: 591-592. https://doi.org/10.1042/bst0110591
|
| [51] |
Fales F (1951) The assimilation and degradation of carbohydrates by yeast cells. J Biol Chem 193: 113-124. https://doi.org/10.1016/S0021-9258(19)52433-4
|
| [52] |
Lowry OH (1951) Protein measurement with the Folin-Phenol reagents. J Biol Chem 193: 265-275. https://doi.org/10.1016/S0021-9258(19)52451-6
|
| [53] |
Shinwari ZK, Tanveer F, Iqrar I (2019) Role of microbes in plant health, disease management, and abiotic stress management. Microbiome in Plant Health and Disease: Challenges and Opportunities . Singapore: Springer 231-250. https://doi.org/10.1007/978-981-13-8495-0_11
|
| [54] | Pundir RK, Jain P (2015) Mechanism of prevention and control of medicinal plant-associated diseases. Microbiome in Plant Health and Disease . Singapore: Springer 231-246. https://doi.org/10.1007/978-981-13-8495-0_11 |
| [55] |
Hassane AMA, Taha TM, Awad MF, et al. (2022) Radical scavenging potency, HPLC profiling and phylogenetic analysis of endophytic fungi isolated from selected medicinal plants of Saudi Arabia. Electron J Biotechnol 58: 37-45. https://doi.org/10.1016/j.ejbt.2022.05.001
|
| [56] |
Lazreg F, Belabid L (2013) First report of Fusarium chlamydosporum causing damping off disease on Aleppo pine in Algeria. Plant Dis 97: 1506. https://doi.org/10.1094/PDIS-02-13-0208-PDN
|
| [57] |
Mohamed H, Awad MF, Shah AM, et al. (2022) Coculturing of Mucor plumbeus and Bacillus subtilis bacterium as an efficient fermentation strategy to enhance fungal lipid and gamma-linolenic acid (GLA) production. Sci Rep 12: 13111. https://doi.org/10.1038/s41598-022-17442-2
|
| [58] |
Mazrou YS, Makhlouf AH, Elbealy ER, et al. (2020) Molecular characterization of phosphate solubilizing fungi Aspergillus niger and its correlation to sustainable agriculture. J Environ Biol 41: 592-599. https://doi.org/10.22438/jeb/41/3/MRN-1298
|
| [59] |
Palmer CM, Alper HS (2019) Expanding the chemical palette of industrial microbes: Metabolic engineering for type III PKS-derived polyketides. Biotechnol J 14: 1700463. https://doi.org/10.1002/biot.201700463
|
| [60] |
Hussain H, Jabeen F, Krohn K, et al. (2015) Antimicrobial activity of two mellein derivatives isolated from an endophytic fungus. Med Chem Res 24: 2111-2114. https://doi.org/10.1007/s00044-014-1250-3
|
| [61] | Mdachi S (2016) Naturally occurring mellein-type 3,4-dihydroisocoumarins and related lactones: Synthetic approaches-a review. Tanz J Sci 42: 23-62. |
| [62] |
Shaaban R, Elnaggar MS, Khalil N, et al. (2023) A comprehensive review on the medicinally valuable endosymbiotic fungi Penicillium chrysogenum. Arch Microbiol 205: 1-28. https://doi.org/10.1007/s00203-023-03580-2
|
| [63] |
Karpova NV, Yaderets VV, Glagoleva EV, et al. (2021) Antifungal activity of the dry biomass of Penicillium chrysogenum f-24-28 and is application in combination with azoxystrobin for efficient crop protection. Agriculture 11: 935. https://doi.org/10.3390/agriculture11100935
|
| [64] |
Sasidharan ATK, Elyas KK (2019) Anti-fungal potential and brine shrimp lethality assay of in vitro raised clones of Celastrus paniculatus. Asian J Biol Sci 12: 877-883. https://doi.org/10.3923/ajbs.2019.877.883
|
| [65] | Saber SM, Youssef MS, Arafa RF, et al. (2016) Mycotoxins production by Aspergillus ostianus Wehmer and using phytochemicals as control agent. J Sci Eng Res 3: 198-213. |
| [66] |
Hajdú Z, Hohmann J, Forgo P, et al. (2007) Diterpenoids and flavonoids from the fruits of Vitex agnus-castus and antioxidant activity of the fruit extracts and their constituents. Phytother Res 21: 391-394. https://doi.org/10.1002/ptr.2021
|
| [67] |
Chowdhury NS, Sohrab MH, Rana MS, et al. (2017) Cytotoxic naphthoquinone and azaanthraquinone derivatives from an endophytic Fusarium solani. J Nat Prod 80: 1173-1177. https://doi.org/10.1021/acs.jnatprod.6b00610
|
| [68] | Rongai D, Pulcini P, Pesce B, et al. (2015) Antifungal activity of some botanical extracts on Fusarium oxysporum. Open Life Sci 10: 409-416. https://doi.org/10.1515/biol-2015-0040 |
| [69] |
El Hadrami A, Adam LR, Daayf F (2011) Biocontrol treatments confer protection against Verticillium dahliae infection of potato by inducing antimicrobial metabolites. Mol Plant Microbe Interact 24: 328-335. https://doi.org/10.1094/MPMI-04-10-0098
|
| [70] | Stanković MS, Stefanović O, Čomić L, et al. (2012) Antimicrobial activity, total phenolic content and flavonoid concentrations of Teucrium species. Cent Eur J Biol 7: 664-671. https://doi.org/10.2478/s11535-012-0048-x |
| [71] |
Vinale F, Nicoletti R, Borrelli F, et al. (2017) Co-culture of plant beneficial microbes as source of bioactive metabolites. Sci Rep 7: 14330. https://doi.org/10.1038/s41598-017-14569-5
|
| [72] |
Peng X, Wang Y, Sun K, et al. (2011) Cerebrosides and 2-pyridone alkaloids from the halotolerant fungus Penicillium chrysogenum grown in a hypersaline medium. J Nat Prod 74: 1298-1302. https://doi.org/10.1021/np1008976
|
| [73] |
Liu Z, Fu B, Duan X, et al. (2022) Effects of cell-cell interactions between A. oryzae and Z. rouxii on morphology and secondary metabolites. LWT 170: 114035. https://doi.org/10.1016/j.lwt.2022.114035
|
| [74] |
Rateb ME, Hallyburton I, Houssen WE, et al. (2013) Induction of diverse secondary metabolites in Aspergillus fumigatus by microbial co-culture. RSC Adv 3: 14444-14450. https://doi.org/10.1039/c3ra42378f
|
| [75] | König CC, Scherlach K, Schroeckh V, et al. (2013) Bacterium induces cryptic meroterpenoid pathway in the pathogenic fungus Aspergillus fumigatus. Chem Biochem 14: 938-942. https://doi.org/10.1002/cbic.201300070 |
| [76] |
Ninomiya A, Urayama S, Hagiwara D (2022) Antibacterial diphenyl ether production induced by co-culture of Aspergillus nidulans and Aspergillus fumigatus. Appl Microbiol Biotechnol 106: 4169-4185. https://doi.org/10.1007/s00253-022-11964-5
|
| [77] |
Wang Y, Glukhov E, He Y, et al. (2022) Secondary metabolite variation and bioactivities of two marine Aspergillus strains in static co-culture investigated by molecular network analysis and multiple database mining based on LC-PDA-MS/MS. Antibiotics 11: 513. https://doi.org/10.3390/antibiotics11040513
|
| [78] |
El-Sayed ASA, Shindia AA, AbouZeid A, et al. (2021) Triggering the biosynthetic machinery of Taxol by Aspergillus flavipes via cocultivation with Bacillus subtilis: Proteomic analyses emphasize the chromatin remodeling upon fungal-bacterial interaction. Environ Sci Pollut Res 28: 39866-39881. https://doi.org/10.1007/s11356-021-13533-1
|
| [79] |
Hoshino S, Onaka H, Abe I (2019) Activation of silent biosynthetic pathways and discovery of novel secondary metabolites in actinomycetes by co-culture with mycolic acid-containing bacteria. J Ind Microbiol Biotechnol 46: 363-374. https://doi.org/10.1007/s10295-018-2100-y
|
| [80] |
Attia MS, Salem MS, Abdelaziz AM (2024) Endophytic fungi Aspergillus spp. reduce fusarial wilt disease severity, enhance growth, metabolism and stimulate the plant defense system in pepper plants. Biomass Conv Bioref 14: 16603-16613. https://doi.org/10.1007/s13399-022-03607-6
|
| [81] |
Hussein MMA, Abo-Elyousr KAM, Hassan MAH, et al. (2018) Induction of defense mechanisms involved in disease resistance of onion blight disease caused by Botrytis allii. Egypt J Biol Pest Control 28: 80. https://doi.org/10.1186/s41938-018-0085-5
|
| [82] | Pessel N, Balmat JF (2008) Principal component analysis for greenhouse modelling. WSEAS Trans Syst 7: 24-30. |
| [83] |
Konishi T (2015) Principal component analysis for designed experiments. BMC Bioinform 16: 1-9. https://doi.org/10.1186/1471-2105-16-S18-S7
|
 microbiol-10-04-044-s001.pdf microbiol-10-04-044-s001.pdf |
 |














Figures(15)

Mohammed M. M. Abdelrahem, Mohamed E. Abouelela, Nageh F. Abo-Dahab, Abdallah M. A. Hassane. Aspergillus-Penicillium co-culture: An investigation of bioagents for controlling Fusarium proliferatum-induced basal rot in onion[J]. AIMS Microbiology, 2024, 10(4): 1024-1051. doi: 10.3934/microbiol.2024044







 DownLoad:
DownLoad: