Citation: Yoram Baram. Asynchronous Segregation of Cortical Circuits and Their Function: A Life-long Role for Synaptic Death[J]. AIMS Neuroscience, 2017, 4(2): 87-101. doi: 10.3934/Neuroscience.2017.2.87

| [1] | Huttenlocher PR (1979) Synaptic density in human frontal cortex. Development changes and effects of age. Brain Res 163: 195–205. |
| [2] |
Huttenlocher PR, De Courten C, Garey LJ, et al. (1982) Synaptogenesis in human visual cortex-evidence for synapse elimination during normal development. Neurosci Lett 33: 247–252. doi: 10.1016/0304-3940(82)90379-2

|
| [3] | Huttenlocher PR, De Courten C (1987) The development of synapses in striate cortex of man. J Neurosci 6: 1–9. |
| [4] |
Eckenhoff MF, Rakic P (1991) A quantative analysis of synaptogenesis in the molecular layer of the dentate gyrus in the resus monkey. Develop Brain Res 64: 129–135. doi: 10.1016/0165-3806(91)90216-6
|
| [5] | Bourgeois JP (1993) Synaptogenesis in the prefrontal cortex of the Macaque. In: B. do Boysson–Bardies (Ed.), Develop neurocog: Speech and face processing in the first year of life. Norwell, MA; Kluwer: 31–39. |
| [6] | Bourgeois JP, Rakic P (1993) Changing of synaptic density in the primary visual cortex of the rhesus monkey from fetal to adult age. J Neurosci 13:2801–2820. |
| [7] |
Rakic P, Bourgeois JP, Goldman-Rakic, et al. (1994) Synaptic development of the cerebral cortex: Implications for learning, memory and mental illness. Prog in Brain Res 102: 227–243. doi: 10.1016/S0079-6123(08)60543-9
|
| [8] |
Innocenti GM (1995) Exuberant development of connections and its possible permissive role in cortical evolution. Trends Neurosci 18: 397–402. doi: 10.1016/0166-2236(95)93936-R
|
| [9] |
Takacs J, Hamori J (1994) Developmental dynamics of Purkinje cells and dendritic spines in rat cerebellar cortex. Neurosci Res 38: 515–530. doi: 10.1002/jnr.490380505
|
| [10] |
Dennis MJ, Yip JW (1978) Formation and elimination of foreign synapses on adult salamander muscle. J Physiol 274: 299–310. doi: 10.1113/jphysiol.1978.sp012148
|
| [11] |
Balice-Gordon RJ, Lichtman JW (1994) Long–term synapse loss induced by focal blockade of postsynaptlc receptors. Nature 372: 519 – 524. doi: 10.1038/372519a0
|
| [12] |
Balice-Gordon RJ, Chua C, Nelson CC, et al. (1993) Gradual loss of synaptic cartels precedes axon withdrawal at developing neuromuscular junctions. Neuron 11: 801–815. doi: 10.1016/0896-6273(93)90110-D
|
| [13] | Vanderhaeghen P, Cheng HJ (2010) Guidance Molecules in Axon Pruning and Cell death. Cold Spring Harbor Perspect in Biol 2: 1–18. |
| [14] |
Atwood HL, Wojtowicz JM (1999) Silent Synapses in Neural Plasticity: Current Evidence. Learn Mem 6: 542–571. doi: 10.1101/lm.6.6.542
|
| [15] | Culican SM, Nelson CC, Lichtman JW (1998) Axon withdrawal during synapse elimination at the neuromuscular junction is accompanied by disassembly of the postsynaptic specialization and withdrawal of schwann cell processes. J Neurosci 18: 4953–4965. |
| [16] |
Chechik G, Meilijson I, Ruppin E (1998) Synaptic pruning in development: a computational account. Neur comp 10: 1759–1777. doi: 10.1162/089976698300017124
|
| [17] | Iglesias J, Eriksson J, Grize F, et al. (2005) Dynamics of pruning in simulated large–scale spiking neural networks. BioSys 79: 11–20. |
| [18] |
Giedd JN, Blumenthal J, Jeffries NO, et al. (1999) Brain development during childhood and adolescence: a longitudinal MRI study. Nat Neurosci 2: 861–863. doi: 10.1038/13158
|
| [19] |
Mechelli A, Crinion JT, Noppeney U, et al. (2004) Structural plasticity in the bilingual brain. Nature 431: 757. doi: 10.1038/431757a
|
| [20] |
Craik F, Bialystok E (2006) Cognition through the lifespan: mechanisms of change. Trends Cogn Sci 10: 131–138. doi: 10.1016/j.tics.2006.01.007
|
| [21] |
Lee YI, Li Y, Mikesh M, et al. (2016) Neuregulin1 displayed on motor axons regulates terminal Schwann cell–mediated synapse elimination at developing neuromuscular junctions. PNAS 113: E479–E487. doi: 10.1073/pnas.1519156113
|
| [22] |
Baram Y (2017) Developmental metaplasticity in neural circuit codes of firing and structure. Neur Net 85: 182–196. doi: 10.1016/j.neunet.2016.09.007
|
| [23] | Lapicque L (1907) Recherches quantitatives sur l'excitation électrique des nerfs traitée comme une polarisation. J Physiol Pathol Gen 9: 620–635. |
| [24] |
Hodgkin A, Huxley AA (1952) Quantitative description of membrane current and its application to conduction and excitation in nerve. J Physiol 117: 500–544. doi: 10.1113/jphysiol.1952.sp004764
|
| [25] | Wilson HR, Cowan JD (1972) Excitatory and Inhibitory Interactions in Localized Populations of Model Neurons. Biophys J 12: 1–24. |
| [26] |
Abbott LF (1994) Decoding neuronal firing and modeling neural networks. Quart Rev Biophys 27: 291–331. doi: 10.1017/S0033583500003024
|
| [27] | Gerstner W (1995)Time structure of the activity in neural network models. Phys Rev E 51: 738–758. |
| [28] |
Baram Y (2013) Global attractor alphabet of neural firing modes. J Neurophys 110: 907–915. doi: 10.1152/jn.00107.2013
|
| [29] |
Granit RD, Kernell D, Shortess GK (1963) Quantitative aspects of repetitive firing of mammalian motoneurons caused by injected currents. J Physiol 168: 911–931. doi: 10.1113/jphysiol.1963.sp007230
|
| [30] |
Connor JA, Stevens CF (1971) Voltage clamp studies of a transient outward membrane current in gastropod neural somata. J Physiol 213: 21–30. doi: 10.1113/jphysiol.1971.sp009365
|
| [31] | Carandini M, Ferster D (2000) Membrane Potential and Firing Rate in Cat Primary Visual Cortex. J Neurosci 20: 470–484. |
| [32] | Bienenstock EL, Cooper LN, Munro PW (1982) Theory for the development of neuron selectivity: orientation specificity and binocular interaction in visual cortex. J Neurosci 2: 32–48. |
| [33] |
Intrator N, Cooper LN (1992) Objective function formulation of the BCM theory of visual cortical plasticity: Statistical connections, stability conditions. Neur Netw 5: 3–17. doi: 10.1016/S0893-6080(05)80003-6
|
| [34] | Cooper LN, Intrator N, Blais BS, et al. (2004) Theory of Cortical Plasticity. New Jersey: World Sciettific. |
| [35] | Hebb DO (1949) The organization of behavior: a neuropsychological theory. New York: Wiley. |
| [36] | Koenigs G (1884) Rescherches sur les integrals de certains equations fonctionnelles. Ann Sci de l'Ecole Normale Super 3: 3–41. |
| [37] | Lemeray EM (1895) Sur les fonctions iteratives et sur une nouvelle fonctions. Assoc Franc pour l'Advence des Sci, Congres Bordeuaux 2: 149–165. |
| [38] |
Knoebel RA (1981) Exponential reiterated. Amer Math Month 88: 235–252. doi: 10.2307/2320546
|
| [39] | Abraham RH, Gardini L, Mira C (1997) Chaos in Discrete Dynamical Systems. Berlin: Springer-Verlag. |
| [40] |
Li TY, Yorke JA (1975) Period three implies chaos. Am Math Month 82: 985–992. doi: 10.2307/2318254
|
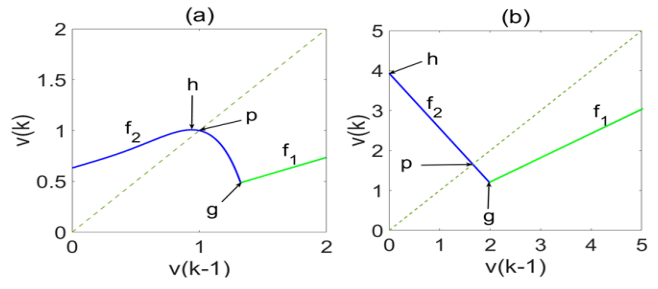
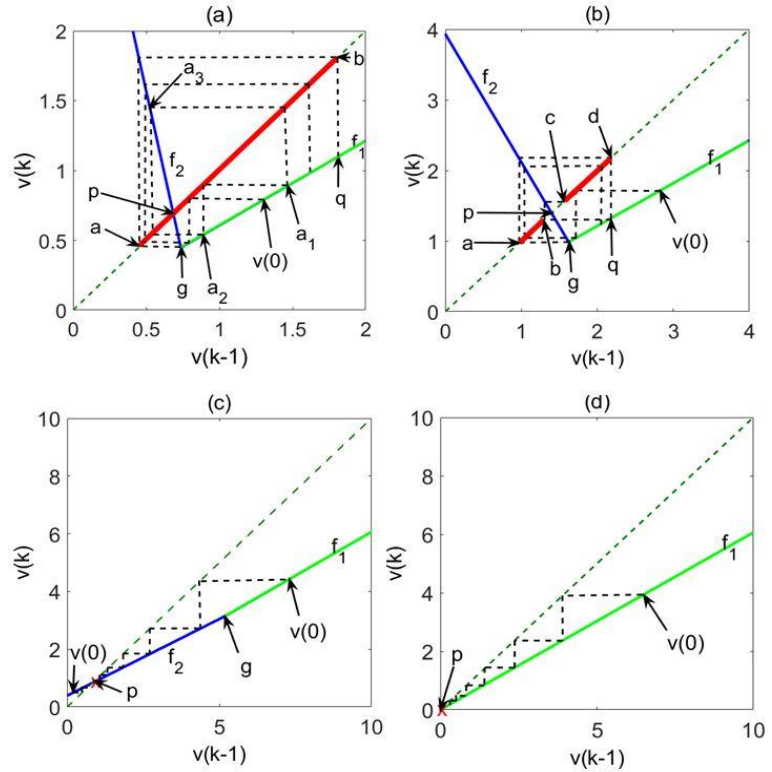
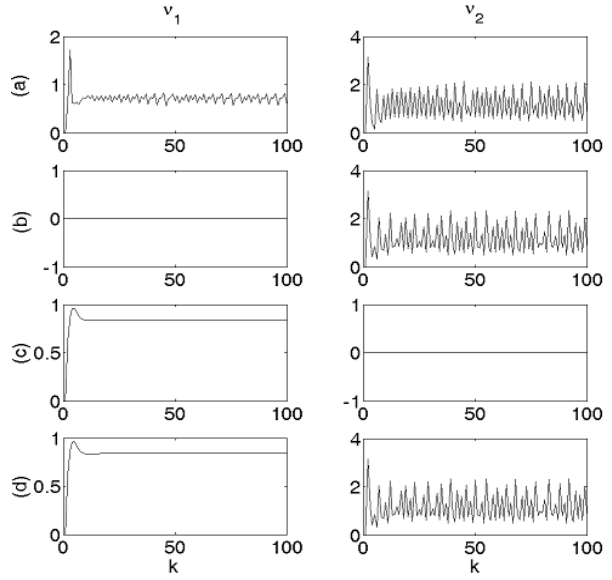
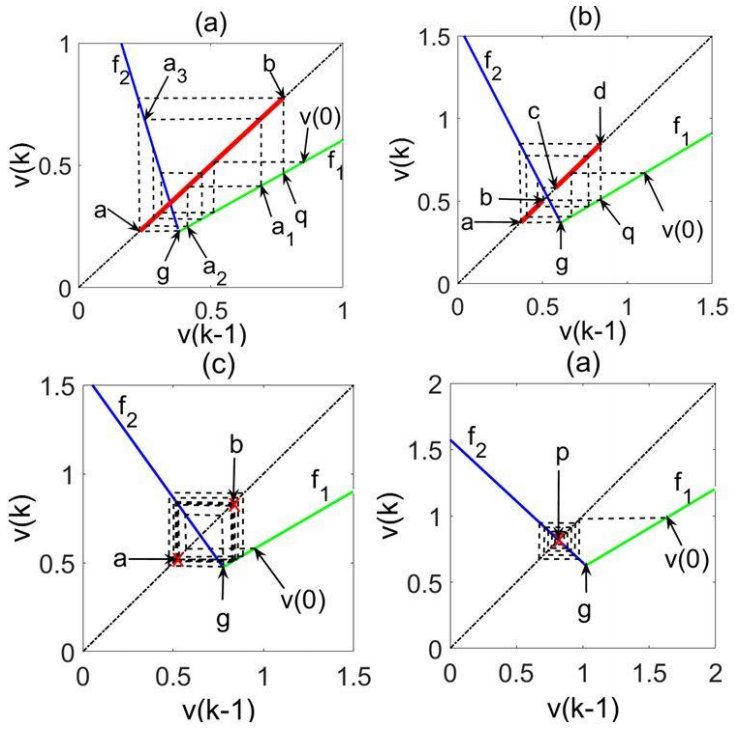
Figures(6)

Yoram Baram. Asynchronous Segregation of Cortical Circuits and Their Function: A Life-long Role for Synaptic Death[J]. AIMS Neuroscience, 2017, 4(2): 87-101. doi: 10.3934/Neuroscience.2017.2.87







 DownLoad:
DownLoad: