Citation: Manjusha Lekshmi, Parvathi Ammini, Jones Adjei, Leslie M. Sanford, Ugina Shrestha, Sanath Kumar, Manuel F. Varela. Modulation of antimicrobial efflux pumps of the major facilitator superfamily in Staphylococcus aureus[J]. AIMS Microbiology, 2018, 4(1): 1-18. doi: 10.3934/microbiol.2018.1.1

| [1] |
Levy SB (1992) Active efflux mechanisms for antimicrobial resistance. Antimicrob Agents Ch 36: 695–703. doi: 10.1128/AAC.36.4.695

|
| [2] |
Higgins CF (2007) Multiple molecular mechanisms for multidrug resistance transporters. Nature 446: 749–757. doi: 10.1038/nature05630
|
| [3] |
Sadee W, Drubbisch V, Amidon GL (1995) Biology of membrane transport proteins. Pharm Res 12: 1823–1837. doi: 10.1023/A:1016211015926
|
| [4] |
West IC (1980) Energy coupling in secondary active transport. BBA-Rev Biomembranes 604: 91–126. doi: 10.1016/0005-2736(80)90586-6
|
| [5] |
Boudker O, Verdon G (2010) Structural perspectives on secondary active transporters. Trends Pharmacol Sci 31: 418–426. doi: 10.1016/j.tips.2010.06.004
|
| [6] |
Kumar S, Varela MF (2012) Biochemistry of bacterial multidrug efflux pumps. Int J Mol Sci 13: 4484–4495. doi: 10.3390/ijms13044484
|
| [7] |
Schindler BD, Kaatz GW (2016) Multidrug efflux pumps of Gram-positive bacteria. Drug Resist Update 27: 1–13. doi: 10.1016/j.drup.2016.04.003
|
| [8] |
Alnaseri H, Arsic B, Schneider JE, et al. (2015) Inducible expression of a resistance-nodulation-division-type efflux pump in Staphylococcus aureus provides resistance to linoleic and arachidonic acids. J Bacteriol 197: 1893–1905. doi: 10.1128/JB.02607-14
|
| [9] |
Chen J, Morita Y, Huda MN, et al. (2002) VmrA, a member of a novel class of Na+-coupled multidrug efflux pumps from Vibrio parahaemolyticus. J Bacteriol 184: 572–576. doi: 10.1128/JB.184.2.572-576.2002
|
| [10] | Evans K, Passador L, Srikumar R, et al. (1998) Influence of the MexAB-OprM multidrug efflux system on quorum sensing in Pseudomonas aeruginosa. J Bacteriol 180: 5443–5447. |
| [11] |
Martinez JL, Sanchez MB, Martinez-Solano L, et al. (2009) Functional role of bacterial multidrug efflux pumps in microbial natural ecosystems. FEMS Microbiol Rev 33: 430–449. doi: 10.1111/j.1574-6976.2008.00157.x
|
| [12] |
Marger MD, Saier MH (1993) A major superfamily of transmembrane facilitators that catalyse uniport, symport and antiport. Trends Biochem Sci 18: 13–20. doi: 10.1016/0968-0004(93)90081-W
|
| [13] | Pao SS, Paulsen IT, Saier MH (1998) Major facilitator superfamily. Microbiol Mol Biol R 62: 1–34. |
| [14] | Paulsen IT, Brown MH, Skurray RA (1996) Proton-dependent multidrug efflux systems. Microbiol Rev 60: 575–608. |
| [15] |
Van Bambeke F, Balzi E, Tulkens PM (2000) Antibiotic efflux pumps. Biochem Pharmacol 60: 457–470. doi: 10.1016/S0006-2952(00)00291-4
|
| [16] | Ranaweera I, Shrestha U, Ranjana KC, et al. (2015) Structural comparison of bacterial multidrug efflux pumps of the major facilitator superfamily. Trends Cell Mol Biol 10: 131–140. |
| [17] |
Murakami S, Yamaguchi A (2003) Multidrug-exporting secondary transporters. Curr Opin Struc Biol 13: 443–452. doi: 10.1016/S0959-440X(03)00109-X
|
| [18] |
Andersen JL, He GX, Kakarla P, et al. (2015) Multidrug efflux pumps from Enterobacteriaceae, Vibrio cholerae and Staphylococcus aureus bacterial food pathogens. Int J Env Res Pub He 12: 1487–1547. doi: 10.3390/ijerph120201487
|
| [19] |
Saier MH, Paulsen IT (2001) Phylogeny of multidrug transporters. Semin Cell Dev Biol 12: 205–213. doi: 10.1006/scdb.2000.0246
|
| [20] |
Putman M, van Veen HW, Konings WN (2000) Molecular properties of bacterial multidrug transporters. Microbiol Mol Biol R 64: 672–693. doi: 10.1128/MMBR.64.4.672-693.2000
|
| [21] | Bay DC, Rommens KL, Turner RJ (2008) Small multidrug resistance proteins: a multidrug transporter family that continues to grow. BBA-Biomembranes 1778: 1814–1838. |
| [22] |
Costa SS, Viveiros M, Amaral L, et al. (2013) Multidrug efflux pumps in Staphylococcus aureus: an update. Open Microbiol J 7: 59–71. doi: 10.2174/1874285801307010059
|
| [23] |
Jang S (2016) Multidrug efflux pumps in Staphylococcus aureus and their clinical implications. J Microbiol 54: 1–8. doi: 10.1007/s12275-016-5159-z
|
| [24] |
McAleese F, Petersen P, Ruzin A, et al. (2005) A novel MATE family efflux pump contributes to the reduced susceptibility of laboratory-derived Staphylococcus aureus mutants to tigecycline. Antimicrob Agents Ch 49: 1865–1871. doi: 10.1128/AAC.49.5.1865-1871.2005
|
| [25] |
Koronakis V, Sharff A, Koronakis E, et al. (2000) Crystal structure of the bacterial membrane protein TolC central to multidrug efflux and protein export. Nature 405: 914–919. doi: 10.1038/35016007
|
| [26] |
Piddock LJ (2006) Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin Microbiol Rev 19: 382–402. doi: 10.1128/CMR.19.2.382-402.2006
|
| [27] | Alvarez-Ortega C, Olivares J, Martinez JL (2013) RND multidrug efflux pumps: what are they good for? Front Microbiol 4: 7. |
| [28] |
Murakami S, Nakashima R, Yamashita E, et al. (2002) Crystal structure of bacterial multidrug efflux transporter AcrB. Nature 419: 587–593. doi: 10.1038/nature01050
|
| [29] |
Nakashima R, Sakurai K, Yamasaki S, et al. (2011) Structures of the multidrug exporter AcrB reveal a proximal multisite drug-binding pocket. Nature 480: 565–569. doi: 10.1038/nature10641
|
| [30] |
Kuroda M, Ohta T, Uchiyama I, et al. (2001) Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet 357: 1225–1240. doi: 10.1016/S0140-6736(00)04403-2
|
| [31] |
Smith KP, Kumar S, Varela MF (2009) Identification, cloning, and functional characterization of EmrD-3, a putative multidrug efflux pump of the major facilitator superfamily from Vibrio cholerae O395. Arch Microbiol 191: 903–911. doi: 10.1007/s00203-009-0521-8
|
| [32] |
Floyd JL, Smith KP, Kumar SH, et al. (2010) LmrS is a multidrug efflux pump of the major facilitator superfamily from Staphylococcus aureus. Antimicrob Agents Ch 54: 5406–5412. doi: 10.1128/AAC.00580-10
|
| [33] |
Ubukata K, Itoh-Yamashita N, Konno M (1989) Cloning and expression of the norA gene for fluoroquinolone resistance in Staphylococcus aureus. Antimicrob Agents Ch 33: 1535–1539. doi: 10.1128/AAC.33.9.1535
|
| [34] |
Truong-Bolduc QC, Dunman PM, Strahilevitz J, et al. (2005) MgrA is a multiple regulator of two new efflux pumps in Staphylococcus aureus. J Bacteriol 187: 2395–2405. doi: 10.1128/JB.187.7.2395-2405.2005
|
| [35] |
Yoshida H, Bogaki M, Nakamura S, et al. (1990) Nucleotide sequence and characterization of the Staphylococcus aureus norA gene, which confers resistance to quinolones. J Bacteriol 172: 6942–6949. doi: 10.1128/jb.172.12.6942-6949.1990
|
| [36] |
Kaatz GW, Seo SM, Ruble CA (1993) Efflux-mediated fluoroquinolone resistance in Staphylococcus aureus. Antimicrob Agents Ch 37: 1086–1094. doi: 10.1128/AAC.37.5.1086
|
| [37] |
Ng EY, Trucksis M, Hooper DC (1994) Quinolone resistance mediated by norA: physiologic characterization and relationship to flqB, a quinolone resistance locus on the Staphylococcus aureus chromosome. Antimicrob Agents Ch 38: 1345–1355. doi: 10.1128/AAC.38.6.1345
|
| [38] |
Ding Y, Onodera Y, Lee JC, et al. (2008) NorB, an efflux pump in Staphylococcus aureus strain MW2, contributes to bacterial fitness in abscesses. J Bacteriol 190: 7123–7129. doi: 10.1128/JB.00655-08
|
| [39] |
Truong-Bolduc QC, Bolduc GR, Okumura R, et al. (2011) Implication of the NorB efflux pump in the adaptation of Staphylococcus aureus to growth at acid pH and in resistance to moxifloxacin. Antimicrob Agents Ch 55: 3214–3219. doi: 10.1128/AAC.00289-11
|
| [40] |
Truong-Bolduc QC, Strahilevitz J, Hooper DC (2006) NorC, a new efflux pump regulated by MgrA of Staphylococcus aureus. Antimicrob Agents Ch 50: 1104–1107. doi: 10.1128/AAC.50.3.1104-1107.2006
|
| [41] |
Paulsen IT, Brown MH, Littlejohn TG, et al. (1996) Multidrug resistance proteins QacA and QacB from Staphylococcus aureus: membrane topology and identification of residues involved in substrate specificity. Proc Natl Acad Sci USA 93: 3630–3635. doi: 10.1073/pnas.93.8.3630
|
| [42] | Tennent JM, Lyon BR, Midgley M, et al. (1989) Physical and biochemical characterization of the qacA gene encoding antiseptic and disinfectant resistance in Staphylococcus aureus. J Gen Microbiol 135: 1–10. |
| [43] |
Rouch DA, Cram DS, Berardino DD, et al. (1990) Efflux-mediated antiseptic resistance gene qacA from Staphylococcus aureus: common ancestry with tetracycline- and sugar-transport proteins. Mol Microbiol 4: 2051–2062. doi: 10.1111/j.1365-2958.1990.tb00565.x
|
| [44] |
Yamada Y, Hideka K, Shiota S, et al. (2006) Gene cloning and characterization of SdrM, a chromosomally-encoded multidrug efflux pump, from Staphylococcus aureus. Biol Pharm Bull 29: 554–556. doi: 10.1248/bpb.29.554
|
| [45] |
Truong-Bolduc QC, Bolduc GR, Medeiros H, et al. (2015) Role of the Tet38 efflux pump in Staphylococcus aureus internalization and survival in epithelial cells. Infect Immun 83: 4362–4372. doi: 10.1128/IAI.00723-15
|
| [46] |
Ginn SL, Brown MH, Skurray RA (2000) The TetA(K) tetracycline/H+ antiporter from Staphylococcus aureus: mutagenesis and functional analysis of motif C. J Bacteriol 182: 1492–1498. doi: 10.1128/JB.182.6.1492-1498.2000
|
| [47] | Krulwich TA, Jin J, Guffanti AA, et al. (2001) Functions of tetracycline efflux proteins that do not involve tetracycline. J Mol Microb Biotechnol 3: 237–246. |
| [48] |
Schwarz S, Fessler AT, Hauschild T, et al. (2011) Plasmid-mediated resistance to protein biosynthesis inhibitors in staphylococci. Ann NY Acad Sci 1241: 82–103. doi: 10.1111/j.1749-6632.2011.06275.x
|
| [49] |
Couto I, Costa SS, Viveiros M, et al. (2008) Efflux-mediated response of Staphylococcus aureus exposed to ethidium bromide. J Antimicrob Chemoth 62: 504–513. doi: 10.1093/jac/dkn217
|
| [50] |
Fluman N, Bibi E (2009) Bacterial multidrug transport through the lens of the major facilitator superfamily. BBA-Proteins Proteom 1794: 738–747. doi: 10.1016/j.bbapap.2008.11.020
|
| [51] | Brown MH, Skurray RA (2001) Staphylococcal multidrug efflux protein QacA. J Mol Microb Biotechnol 3: 163–170. |
| [52] |
Ho J, Branley J (2012) Prevalence of antiseptic resistance genes qacA/B and specific sequence types of methicillin-resistant Staphylococcus aureus in the era of hand hygiene. J Antimicrob Chemoth 67: 1549–1550. doi: 10.1093/jac/dks035
|
| [53] |
Truong-Bolduc QC, Villet RA, Estabrooks ZA, et al. (2014) Native efflux pumps contribute resistance to antimicrobials of skin and the ability of Staphylococcus aureus to colonize skin. J Infect Dis 209: 1485–1493. doi: 10.1093/infdis/jit660
|
| [54] |
Handzlik J, Matys A, Kiec-Kononowicz K (2013) Recent advances in multi-drug resistance (MDR) efflux pump inhibitors of Gram-positive bacteria S. aureus. Antibiotics 2: 28–45. doi: 10.3390/antibiotics2010028
|
| [55] |
Li XZ, Nikaido H (2009) Efflux-mediated drug resistance in bacteria: an update. Drugs 69: 1555–1623. doi: 10.2165/11317030-000000000-00000
|
| [56] |
Huang J, O'Toole PW, Shen W, et al. (2004) Novel chromosomally encoded multidrug efflux transporter MdeA in Staphylococcus aureus. Antimicrob Agents Ch 48: 909–917. doi: 10.1128/AAC.48.3.909-917.2004
|
| [57] |
Yamada Y, Shiota S, Mizushima T, et al. (2006) Functional gene cloning and characterization of MdeA, a multidrug efflux pump from Staphylococcus aureus. Biol Pharm Bull 29: 801–804. doi: 10.1248/bpb.29.801
|
| [58] |
Grkovic S, Brown MH, Roberts NJ, et al. (1998) QacR is a repressor protein that regulates expression of the Staphylococcus aureus multidrug efflux pump QacA. J Biol Chem 273: 18665–18673. doi: 10.1074/jbc.273.29.18665
|
| [59] |
DeMarco CE, Cushing LA, Frempong-Manso E, et al. (2007) Efflux-related resistance to norfloxacin, dyes, and biocides in bloodstream isolates of Staphylococcus aureus. Antimicrob Agents Ch 51: 3235–3239. doi: 10.1128/AAC.00430-07
|
| [60] | Heytler PG (1963) Uncoupling of oxidative phosphorylation by carbonyl cyanide phenylhydrazones. I. Some characteristics of m-Cl-CCP action on mitochondria and chloroplasts. Biochemistry 2: 357–361. |
| [61] |
Jones I, Midgley M (1985) Expression of a plasmid borne ethidium resistance determinant from Staphylococcus in Escherichia coli: evidence for an efflux system. FEMS Microbiol Lett 28: 355–358. doi: 10.1111/j.1574-6968.1985.tb00820.x
|
| [62] | Mitchell BA, Paulsen IT, Brown MH, et al. (1999) Bioenergetics of the staphylococcal multidrug export protein QacA. Identification of distinct binding sites for monovalent and divalent cations. J Biol Chem 274: 3541–3548. |
| [63] | Littlejohn TG, Paulsen IT, Gillespie MT, et al. (1992) Substrate specificity and energetics of antiseptic and disinfectant resistance in Staphylococcus aureus. FEMS Microbiol Lett 74: 259–265. |
| [64] | Mo XN, Li JG, Tang YC, et al. (2007) The action of active efflux system on multi-drug resistance in methicillin resistant Staphylococcus aureus. Zhonghua Jie He He Hu Xi Za Zhi 30: 40–43. |
| [65] |
Rothstein DM, McGlynn M, Bernan V, et al. (1993) Detection of tetracyclines and efflux pump inhibitors. Antimicrob Agents Ch 37: 1624–1629. doi: 10.1128/AAC.37.8.1624
|
| [66] | Nelson ML, Levy SB (1999) Reversal of tetracycline resistance mediated by different bacterial tetracycline resistance determinants by an inhibitor of the Tet(B) antiport protein. Antimicrob Agents Ch 43: 1719–1724. |
| [67] |
Roccaro AS, Blanco AR, Giuliano F, et al. (2004) Epigallocatechin-gallate enhances the activity of tetracycline in staphylococci by inhibiting its efflux from bacterial cells. Antimicrob Agents Ch 48: 1968–1973. doi: 10.1128/AAC.48.6.1968-1973.2004
|
| [68] | de Araujo RSA, Barbosa-Filho JM, Scotti MT, et al. (2016) Modulation of drug resistance in Staphylococcus aureus with coumarin derivatives. Scientifica 2016: 1–6. |
| [69] | Tintino SR, Morais-Tintino CD, Campina FF, et al. (2016) Action of cholecalciferol and alpha-tocopherol on Staphylococcus aureus efflux pumps. EXCLI J 15: 315–322. |
| [70] |
Monte J, Abreu AC, Borges A, et al. (2014) Antimicrobial activity of selected phytochemicals against Escherichia coli and Staphylococcus aureus and their biofilms. Pathogens 3: 473–498. doi: 10.3390/pathogens3020473
|
| [71] | Limaverde PW, Campina FF, da Cunha FA, et al. (2017) Inhibition of the TetK efflux-pump by the essential oil of Chenopodium ambrosioides L. and alpha-terpinene against Staphylococcus aureus IS-58. Food Chem Toxicol 109: 957–961. |
| [72] |
Chovanova R, Mezovska J, Vaverkova S, et al. (2015) The inhibition the Tet(K) efflux pump of tetracycline resistant Staphylococcus epidermidis by essential oils from three Salvia species. Lett Appl Microbiol 61: 58–62. doi: 10.1111/lam.12424
|
| [73] | Varela MF, Andersen JL, Ranjana KC, et al. (2017) Bacterial resistance mechanisms and inhibitors of multidrug efflux pumps belonging to the major facilitator superfamily of solute transport systems, In: Atta-ur-Rahman, Choudhary MI, Editors, Frontiers in Anti-Infective Drug Discovery, Bentham Science Publishers, 109–131. |
| [74] |
Kumar S, He G, Kakarla P, et al. (2016) Bacterial multidrug efflux pumps of the major facilitator superfamily as targets for modulation. Infect Disord Drug Targets 16: 28–43. doi: 10.2174/1871526516666160407113848
|
| [75] | Kumar S, Mukherjee MM, Varela MF (2013) Modulation of bacterial multidrug resistance efflux pumps of the major facilitator superfamily. Int J Bacteriol 2013: 1–15. |
| [76] |
Pervaiz A, Khan R, Anwar F, et al. (2016) Alkaloids: An emerging antibacterial modality against methicillin resistant Staphylococcus aureus. Curr Pharm Design 22: 4420–4429. doi: 10.2174/1381612822999160629115627
|
| [77] |
Wang Y, Venter H, Ma S (2016) Efflux pump inhibitors: A novel approach to combat efflux-mediated drug resistance in bacteria. Curr Drug Targets 17: 702–719. doi: 10.2174/1389450116666151001103948
|
| [78] |
Schindler BD, Jacinto P, Kaatz GW (2013) Inhibition of drug efflux pumps in Staphylococcus aureus: current status of potentiating existing antibiotics. Future Microbiol 8: 491–507. doi: 10.2217/fmb.13.16
|
| [79] |
Zhang L, Ma S (2010) Efflux pump inhibitors: a strategy to combat P-glycoprotein and the NorA multidrug resistance pump. Chem Med Chem 5: 811–822. doi: 10.1002/cmdc.201000006
|
| [80] |
Lemieux MJ, Song J, Kim MJ, et al. (2003) Three-dimensional crystallization of the Escherichia coli glycerol-3-phosphate transporter: a member of the major facilitator superfamily. Protein Sci 12: 2748–2756. doi: 10.1110/ps.03276603
|
| [81] |
Bhaskar BV, Babu TM, Reddy NV, et al. (2016) Homology modeling, molecular dynamics, and virtual screening of NorA efflux pump inhibitors of Staphylococcus aureus. Drug Des Dev Ther 10: 3237–3252. doi: 10.2147/DDDT.S113556
|
| [82] | Kumar S, Ranjana KC, Sanford LM, et al. (2016) Structural and functional roles of two evolutionarily conserved amino acid sequence motifs within solute transporters of the major facilitator superfamily. Trends Cell Mol Biol 11: 41–53. |
| [83] | Kakarla P, Ranjana KC, Shrestha U, et al. (2017) Functional roles of highly conserved amino acid sequence motifs A and C in solute transporters of the major facilitator superfamily, In: Arora G, Sajid A, Kalia VC, Editors, Drug Resistance in Bacteria, Fungi, Malaria, and Cancer, Springer International Publishing, 111–140. |
| [84] |
Varela MF, Griffith JK (1993) Nucleotide and deduced protein sequences of the class D tetracycline resistance determinant: relationship to other antimicrobial transport proteins. Antimicrob Agents Ch 37: 1253–1258. doi: 10.1128/AAC.37.6.1253
|
| [85] |
Varela MF, Sansom CE, Griffith JK (1995) Mutational analysis and molecular modelling of an amino acid sequence motif conserved in antiporters but not symporters in a transporter superfamily. Mol Membr Biol 12: 313–319. doi: 10.3109/09687689509072433
|
| [86] |
Yaffe D, Radestock S, Shuster Y, et al. (2013) Identification of molecular hinge points mediating alternating access in the vesicular monoamine transporter VMAT2. Proc Natl Acad Sci USA 110: E1332–E1341. doi: 10.1073/pnas.1220497110
|
| [87] |
Luo J, Parsons SM (2010) Conformational propensities of peptides mimicking transmembrane helix 5 and motif C in wild-type and mutant vesicular acetylcholine transporters. ACS Chem Neurosci 1: 381–390. doi: 10.1021/cn900033s
|
| [88] |
Fontaine F, Hequet A, Voisin-Chiret AS, et al. (2014) First identification of boronic species as novel potential inhibitors of the Staphylococcus aureus NorA efflux pump. J Med Chem 57: 2536–2548. doi: 10.1021/jm401808n
|
| [89] |
Fontaine F, Hequet A, Voisin-Chiret AS, et al. (2015) Boronic species as promising inhibitors of the Staphylococcus aureus NorA efflux pump: study of 6-substituted pyridine-3-boronic acid derivatives. Eur J Med Chem 95: 185–198. doi: 10.1016/j.ejmech.2015.02.056
|
| [90] |
Bharate JB, Singh S, Wani A, et al. (2015) Discovery of 4-acetyl-3-(4-fluorophenyl)-1-(p-tolyl)-5-methylpyrrole as a dual inhibitor of human P-glycoprotein and Staphylococcus aureus Nor A efflux pump. Org Biomol Chem 13: 5424–5431. doi: 10.1039/C5OB00246J
|
| [91] |
Singh S, Kalia NP, Joshi P, et al. (2017) Boeravinone B, A novel dual inhibitor of NorA bacterial efflux pump of Staphylococcus aureus and human P-glycoprotein, reduces the biofilm formation and intracellular invasion of bacteria. Front Microbiol 8: 1868. doi: 10.3389/fmicb.2017.01868
|
| [92] |
Lowrence RC, Raman T, Makala HV, et al. (2016) Dithiazole thione derivative as competitive NorA efflux pump inhibitor to curtail multi drug resistant clinical isolate of MRSA in a zebrafish infection model. Appl Microbiol Biot 100: 9265–9281. doi: 10.1007/s00253-016-7759-2
|
| [93] |
Coelho ML, Ferreira JHL, de Siqueira Junior JP, et al. (2016) Inhibition of the NorA multi-drug transporter by oxygenated monoterpenes. Microb Pathogenesis 99: 173–177. doi: 10.1016/j.micpath.2016.08.026
|
| [94] |
Costa LM, de Macedo EV, Oliveira FA, et al. (2016) Inhibition of the NorA efflux pump of Staphylococcus aureus by synthetic riparins. J Appl Microbiol 121: 1312–1322. doi: 10.1111/jam.13258
|
| [95] |
Tintino SR, Oliveira-Tintino CD, Campina FF, et al. (2016) Evaluation of the tannic acid inhibitory effect against the NorA efflux pump of Staphylococcus aureus. Microb Pathogenesis 97: 9–13. doi: 10.1016/j.micpath.2016.04.003
|
| [96] |
Wani NA, Singh S, Farooq S, et al. (2016) Amino acid amides of piperic acid (PA) and 4-ethylpiperic acid (EPA) as NorA efflux pump inhibitors of Staphylococcus aureus. Bioorg Med Chem Lett 26: 4174–4178. doi: 10.1016/j.bmcl.2016.07.062
|
| [97] | Felicetti T, Cannalire R, Burali MS, et al. (2017) Searching for novel inhibitors of the S. aureus NorA efflux pump: Synthesis and biological evaluation of the 3-Phenyl-1,4-benzothiazine analogues. Chem Med Chem 12: 1293–1302. |
| [98] | Carotti A, Ianni F, Sabatini S, et al. (2016) The "racemic approach" in the evaluation of the enantiomeric NorA efflux pump inhibition activity of 2-phenylquinoline derivatives. J Pharmaceut Biomed 129: 182–189. |
| [99] | Diniz-Silva HT, Magnani M, de Siqueira S, et al. (2016) Fruit flavonoids as modulators of norfloxacin resistance in Staphylococcus aureus that overexpresses norA. LWT-Food Sci Technol 85: 324–326. |
| [100] |
Lepri S, Buonerba F, Goracci L, et al. (2016) Indole-based weapons to fight antibiotic resistance: A structure-activity relationship study. J Med Chem 59: 867–891. doi: 10.1021/acs.jmedchem.5b01219
|
| [101] |
Lopes LAA, dos Santos Rodrigues JB, Magnani M, et al. (2017) Inhibitory effects of flavonoids on biofilm formation by Staphylococcus aureus that overexpresses efflux protein genes. Microb Pathogenesis 107: 193–197. doi: 10.1016/j.micpath.2017.03.033
|
| [102] | Salaheen S, Peng M, Joo J, et al. (2017) Eradication and sensitization of methicillin resistant Staphylococcus aureus to methicillin with bioactive extracts of Berry Pomace. Front Microbiol 8: 253. |
| [103] |
Astolfi A, Felicetti T, Iraci N, et al. (2017) Pharmacophore-based repositioning of approved drugs as novel Staphylococcus aureus NorA efflux pump inhibitors. J Med Chem 60: 1598–1604. doi: 10.1021/acs.jmedchem.6b01439
|
| [104] |
Buonerba F, Lepri S, Goracci L, et al. (2017) Improved potency of indole-based NorA efflux pump inhibitors: From serendipity toward rational design and development. J Med Chem 60: 517–523. doi: 10.1021/acs.jmedchem.6b01281
|
| [105] |
Mirza ZM, Kumar A, Kalia NP, et al. (2011) Piperine as an inhibitor of the MdeA efflux pump of Staphylococcus aureus. J Med Microbiol 60: 1472–1478. doi: 10.1099/jmm.0.033167-0
|
| [106] |
Huet AA, Raygada JL, Mendiratta K, et al. (2008) Multidrug efflux pump overexpression in Staphylococcus aureus after single and multiple in vitro exposures to biocides and dyes. Microbiology 154: 3144–3153. doi: 10.1099/mic.0.2008/021188-0
|
| [107] |
Poole K (2005) Efflux-mediated antimicrobial resistance. J Antimicrob Chemoth 56: 20–51. doi: 10.1093/jac/dki171
|
| [108] |
Luna VA, Heiken M, Judge K, et al. (2002) Distribution of mef(A) in gram-positive bacteria from healthy Portuguese children. Antimicrob Agents Ch 46: 2513–2517. doi: 10.1128/AAC.46.8.2513-2517.2002
|
| [109] |
Kehrenberg C, Schwarz S (2004) fexA, a novel Staphylococcus lentus gene encoding resistance to florfenicol and chloramphenicol. Antimicrob Agents Ch 48: 615–618. doi: 10.1128/AAC.48.2.615-618.2004
|
| [110] |
Kehrenberg C, Schwarz S (2006) Distribution of florfenicol resistance genes fexA and cfr among chloramphenicol-resistant Staphylococcus isolates. Antimicrob Agents Ch 50: 1156–1163. doi: 10.1128/AAC.50.4.1156-1163.2006
|
| [111] |
Kakarla P, Floyd J, Mukherjee M, et al. (2017) Inhibition of the multidrug efflux pump LmrS from Staphylococcus aureus by cumin spice Cuminum cyminum. Arch Microbiol 199: 465–474. doi: 10.1007/s00203-016-1314-5
|
| [112] | Truong-Bolduc QC, Wang Y, Chen C, et al. (2017) Transcriptional regulator TetR21 controls the expression of the Staphylococcus aureus LmrS Efflux pump. Antimicrob Agents Ch 61: e00649-17. |
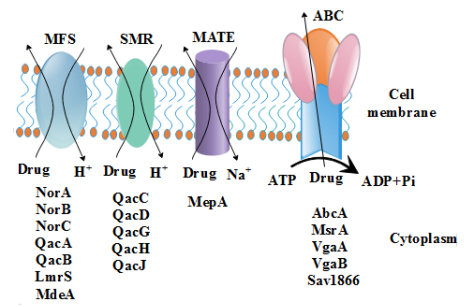
Figures(1) / Tables(1)

Manjusha Lekshmi, Parvathi Ammini, Jones Adjei, Leslie M. Sanford, Ugina Shrestha, Sanath Kumar, Manuel F. Varela. Modulation of antimicrobial efflux pumps of the major facilitator superfamily in Staphylococcus aureus[J]. AIMS Microbiology, 2018, 4(1): 1-18. doi: 10.3934/microbiol.2018.1.1







 DownLoad:
DownLoad: