Technological advances in imaging the human brain help us map and understand the intricacies of cerebral connectivity. Current techniques and specific imaging sequences, however, do come with limitations. Image resolution, variability of techniques and interpretation of images across institutions are just a few concerns. In the setting of high-grade gliomas, understanding how these pathways are affected during tumor growth, surgical resection, and in the brain plasticity presents an even greater challenge. Clinical symptoms, tumor growth, and intraoperative electrical stimulation are important peri-operative considerations to assist in determining neuronal re-wiring and establish a basis of anatomic and functional correlation. The application of functional mapping coupled with the understanding of the natural history of gliomas and implications of neural plasticity, is critical in achieving the goals of maximal tumor resection while minimizing post operative deficits and improving quality of life.
Citation: Sanjay Konakondla, Steven A. Toms. Cerebral Connectivity and High-grade Gliomas: Evolving Concepts of Eloquent Brain in Surgery for Glioma[J]. AIMS Medical Science, 2017, 4(1): 52-70. doi: 10.3934/medsci.2017.1.52
Technological advances in imaging the human brain help us map and understand the intricacies of cerebral connectivity. Current techniques and specific imaging sequences, however, do come with limitations. Image resolution, variability of techniques and interpretation of images across institutions are just a few concerns. In the setting of high-grade gliomas, understanding how these pathways are affected during tumor growth, surgical resection, and in the brain plasticity presents an even greater challenge. Clinical symptoms, tumor growth, and intraoperative electrical stimulation are important peri-operative considerations to assist in determining neuronal re-wiring and establish a basis of anatomic and functional correlation. The application of functional mapping coupled with the understanding of the natural history of gliomas and implications of neural plasticity, is critical in achieving the goals of maximal tumor resection while minimizing post operative deficits and improving quality of life.

| [1] | Duffau H (2015) A two-level model of interindividual anatomo-functional variability of the brain and its implications for neurosurgery. Cortex 1–11. |
| [2] |
Vassal F, Schneider F, Sontheimer A, et al. (2013) Intraoperative visualisation of language fascicles by diffusion tensor imaging-based tractography in glioma surgery. Acta Neurochir (Wien) 155: 437–448. doi: 10.1007/s00701-012-1580-1

|
| [3] |
Roux FE, Boulanouar K, Lotterie JA, et al. (2003) Language functional magnetic resonance imaging in preoperative assessment of language areas: Correlation with direct cortical stimulation. Neurosurgery 52: 1335–1347. doi: 10.1227/01.NEU.0000064803.05077.40
|
| [4] |
Guillevin R, Herpe G, Verdier M, et al. (2014) Low-grade gliomas: The challenges of imaging. Diagn Interv Imaging 95: 957–963. doi: 10.1016/j.diii.2014.07.005
|
| [5] |
Nimsky C, Ganslandt O, Hastreiter P, et al. (2005) Preoperative and intraoperative diffusion tensor imaging-based fiber tracking in glioma surgery. Neurosurgery 56: 130–137. doi: 10.1227/01.NEU.0000144842.18771.30
|
| [6] |
Altman DA, Atkinson DS, Brat DJ (2007) Best cases from the AFIP: glioblastoma multiforme. Radiographics 27: 883–888. doi: 10.1148/rg.273065138
|
| [7] |
Upadhyay N, Waldman AD (2011) Conventional MRI evaluation of gliomas. Br J Radiol 84: 107–111. doi: 10.1259/bjr/65711810
|
| [8] |
Ostrom QT, Gittleman H, Chen Y, et al. (2015) CBTRUS Statistical Report: Primary Brain and Central Nervous System Tumors Diagnosed in the United States in 2008–2012. Neuro Oncol 17: iv1–iv62. doi: 10.1093/neuonc/nov189
|
| [9] |
Glasser MF, Coalson TS, Robinson EC, et al. (2016) A multi-modal parcellation of human cerebral cortex. Nature 536: 171–178. doi: 10.1038/nature18933
|
| [10] |
Fernández Coello A, Moritz-Gasser S, Martino J, et al. (2013) Selection of intraoperative tasks for awake mapping based on relationships between tumor location and functional networks. J Neurosurg 119: 1380–1394. doi: 10.3171/2013.6.JNS122470
|
| [11] |
Duffau H (2014) The huge plastic potential of adult brain and the role of connectomics: New insights provided by serial mappings in glioma surgery. Cortex 58: 325–337. doi: 10.1016/j.cortex.2013.08.005
|
| [12] |
Duffau H, Moritz-Gasser S, Mandonnet E (2014) A re-examination of neural basis of language processing: Proposal of a dynamic hodotopical model from data provided by brain stimulation mapping during picture naming. Brain Lang 131: 1–10. doi: 10.1016/j.bandl.2013.05.011
|
| [13] |
Duffau H (2005) Lessons from brain mapping in surgery for low-grade glioma: Insights into associations between tumour and brain plasticity. Lancet Neurol 4: 476–486. doi: 10.1016/S1474-4422(05)70140-X
|
| [14] |
Duffau H (2013) A plea to pay more attention on anatomo-functional connectivity in surgical management of brain cavernomas. World Neurosurg 80: e221–e223. doi: 10.1016/j.wneu.2012.10.025
|
| [15] | Sawaya R, Hammoud M, Schoppa D, et al. (1998) Neurosurgical Outcomes in a Modern Series of 400 Craniotomies for Treatment of Parenchymal Tumors. Neurosurgery 42. |
| [16] |
Lacroix M, Abi-Said D, Fourney DR, et al. (2001) A multivariate analysis of 416 patients with glioblastoma multiforme: prognosis, extent of resection, and survival. J Neurosurg 95: 190–198. doi: 10.3171/jns.2001.95.2.0190
|
| [17] |
Marko NF, Weil RJ, Schroeder JL, et al. (2014) Extent of resection of glioblastoma revisited: Personalized survival modeling facilitates more accurate survival prediction and supports a maximum-safe-resection approach to surgery. J Clin Oncol 32: 774–782. doi: 10.1200/JCO.2013.51.8886
|
| [18] |
Stupp R, Taillibert S, Kanner AA, et al. (2015) Maintenance Therapy With Tumor-Treating Fields Plus Temozolomide vs Temozolomide Alone for Glioblastoma: A Randomized Clinical Trial. JAMA 314: 2535–2543. doi: 10.1001/jama.2015.16669
|
| [19] |
Liau LM, Prins RM, Kiertscher SM, et al. (2005) Dendritic cell vaccination in glioblastoma patients induces systemic and intracranial T-cell responses modulated by the local central nervous system tumor microenvironment. Clin Cancer Res 11: 5515–5525. doi: 10.1158/1078-0432.CCR-05-0464
|
| [20] |
Berger A (2002) Magnetic resonance imaging. BMJ Br Med J 324: 35. doi: 10.1136/bmj.324.7328.35
|
| [21] |
Gore JCJC (2003) Principles and practice of functional MRI of the human brain. J Clin Invest 112: 4–9. doi: 10.1172/JCI200319010
|
| [22] |
Stadlbauer A, Nimsky C, Buslei R, et al. (2007) Diffusion tensor imaging and optimized fiber tracking in glioma patients: Histopathologic evaluation of tumor-invaded white matter structures. Neuroimage 34: 949–956. doi: 10.1016/j.neuroimage.2006.08.051
|
| [23] |
Kuhnt D, Bauer MHA, Sommer J, et al. (2013) Optic Radiation Fiber Tractography in Glioma Patients Based on High Angular Resolution Diffusion Imaging with Compressed Sensing Compared with Diffusion Tensor Imaging-Initial Experience. Plos One 8: e70973. doi: 10.1371/journal.pone.0070973
|
| [24] |
Price CJ (2012) NeuroImage A review and synthesis of the fi rst 20 years of PET and fMRI studies of heard speech , spoken language and reading. Neuroimage 62: 816–847. doi: 10.1016/j.neuroimage.2012.04.062
|
| [25] | Ueno T, Lambon Ralph MA (2013) The roles of the 'ventral' semantic and 'dorsal' pathways in conduite d'approche: a neuroanatomically-constrained computational modeling investigation. Front Hum Neurosci 7: 422. |
| [26] |
Saur D, Kreher BW, Schnell S, et al. (2008) Ventral and dorsal pathways for language. Proc Natl Acad Sci U S A 105: 18035–18040. doi: 10.1073/pnas.0805234105
|
| [27] |
Huth AG, Heer WA De, Griffiths TL, et al. (2016) Natural speech reveals the semantic maps that tile human cerebral cortex. Nature 532: 453–458. doi: 10.1038/nature17637
|
| [28] |
Tzourio-Mazoyer N, Joliot M, Marie D, et al. (2016) Variation in homotopic areas' activity and inter-hemispheric intrinsic connectivity with type of language lateralization: an FMRI study of covert sentence generation in 297 healthy volunteers. Brain Struct Funct 221: 2735–2753. doi: 10.1007/s00429-015-1068-x
|
| [29] | Rech F, Herbet G, Moritz-Gasser S, et al. (2015) Somatotopic organization of the white matter tracts underpinning motor control in humans: an electrical stimulation study. Brain Struct Funct 221: 3743–3753. |
| [30] |
Bello L, Gambini A, Castellano A, et al. (2008) Motor and language DTI Fiber Tracking combined with intraoperative subcortical mapping for surgical removal of gliomas. Neuroimage 39: 369–382. doi: 10.1016/j.neuroimage.2007.08.031
|
| [31] | Kalani MYS, Kalani M a, Gwinn R, et al. (2009) Embryological development of the human insula and its implications for the spread and resection of insular gliomas. Neurosurg Focus 27: E2. |
| [32] | Michaud K, Duffau H (2016) Surgery of insular and paralimbic diffuse low-grade gliomas: technical considerations. J Neurooncol 1–10. |
| [33] |
Oppenlander ME, Wolf AB, Snyder LA, et al. (2014) An extent of resection threshold for recurrent glioblastoma and its risk for neurological morbidity. J Neurosurg 120: 846–853. doi: 10.3171/2013.12.JNS13184
|
| [34] |
Stupp R, Mason WP, van den Bent MJ, et al. (2005) Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med 352: 987–996. doi: 10.1056/NEJMoa043330
|
| [35] |
Lacroix M, Toms SA (2014) Maximum safe resection of glioblastoma multiforme. J Clin Oncol 32: 727–728. doi: 10.1200/JCO.2013.53.2788
|
| [36] |
Brown PD, Maurer MJ, Rummans TA, et al. (2005) A prospective study of quality of life in adults with newly diagnosed high-grade gliomas: The impact of the extent of resection on quality of life and survival. Neurosurgery 57: 495–503. doi: 10.1227/01.NEU.0000170562.25335.C7
|
| [37] |
Jakola AS, Unsgård G, Solheim O (2011) Quality of life in patients with intracranial gliomas: the impact of modern image-guided surgery. J Neurosurg 114: 1622–1630. doi: 10.3171/2011.1.JNS101657
|
| [38] |
Le Mercier M, Hastir D, Moles Lopez X, et al. (2012) A Simplified Approach for the Molecular Classification of Glioblastomas. PLoS One 7: e45475. doi: 10.1371/journal.pone.0045475
|
| [39] |
Duffau H, Taillandier L (2015) New concepts in the management of diffuse low-grade glioma: Proposal of a multistage and individualized therapeutic approach. Neuro Oncol 17: 332–342. doi: 10.1093/neuonc/nov204.68
|
| [40] |
Duffau H (2013) A new philosophy in surgery for diffuse low-grade glioma (DLGG): Oncological and functional outcomes. Neurochirurgie 59: 2–8. doi: 10.1016/j.neuchi.2012.11.001
|
| [41] |
Pollard SM, Conti L, Sun Y, et al. (2006) Adherent neural stem (NS) cells from fetal and adult forebrain. Cereb Cortex 16: i112–i120. doi: 10.1093/cercor/bhj167
|
| [42] |
Aboody K, Capela A, Niazi N, et al. (2011) Translating Stem Cell Studies to the Clinic for CNS Repair: Current State of the Art and the Need for a Rosetta Stone. Neuron 70: 597–613. doi: 10.1016/j.neuron.2011.05.007
|
| [43] |
Ihrie RA, Álvarez-Buylla A (2011) Lake-Front Property: A Unique Germinal Niche by the Lateral Ventricles of the Adult Brain. Neuron 70: 674–686. doi: 10.1016/j.neuron.2011.05.004
|
| [44] |
Achanta P, Sedora Roman NI, et al. (2010) Gliomagenesis and the use of neural stem cells in brain tumor treatment. Anticancer Agents Med Chem 10: 121–130. doi: 10.2174/187152010790909290
|
| [45] |
Reya T, Morrison SJ, Clarke MF, et al. (2001) Stem cells, cancer, and cancer stem cells. Nature 414: 105–111. doi: 10.1038/35102167
|
| [46] |
Beaulieu C (2002) The basis of anisotropic water diffusion in the nervous system-A technical review. NMR Biomed 15: 435–455. doi: 10.1002/nbm.782
|
| [47] | Ojemann JG, Neil JM, MacLeod a M, et al. (1998) Increased functional vascular response in the region of a glioma. J Cereb Blood Flow Metab 18: 148–153. |
| [48] |
Ojemann JG, Miller JW, Silbergeld DL (1996) Preserved Function in Brain Invaded by Tumor. Neurosurgery 39: 253–259. doi: 10.1097/00006123-199608000-00003
|
| [49] | Bello L, Castellano A, Fava E, et al. (2010) Intraoperative use of diffusion tensor imaging fiber tractography and subcortical mapping for resection of gliomas: technical considerations. Neurosurg Focus 28: E6. |
| [50] |
Morita K, Matsuzawa H, Fujii Y, et al. (2005) Diffusion tensor analysis of peritumoral edema using lambda chart analysis indicative of the heterogeneity of the microstructure within edema. J Neurosurg 102: 336–341. doi: 10.3171/jns.2005.102.2.0336
|
| [51] |
Kinoshita M, Yamada K, Hashimoto N, et al. (2005) Fiber-tracking does not accurately estimate size of fiber bundle in pathological condition: initial neurosurgical experience using neuronavigation and subcortical white matter stimulation. Neuroimage 25: 424–429. doi: 10.1016/j.neuroimage.2004.07.076
|
| [52] |
Giese A, Loo MA, Rief MD, et al. (1995) Substrates for astrocytoma invasion. Neurosurgery 37: 294–301. doi: 10.1227/00006123-199508000-00015
|
| [53] | Giese A, Loo MA, Tran N, et al. (1996) Dichotomy of astrocytoma migration and proliferation. Int J cancer 67: 275–282. |
| [54] |
Giese A, Westphal M (1996) Glioma invasion in the central nervous system. Neurosurgery 39: 235–252. doi: 10.1097/00006123-199608000-00001
|
| [55] |
Oliveira R, Christov C, Guillamo JS, et al. (2005) Contribution of gap junctional communication between tumor cells and astroglia to the invasion of the brain parenchyma by human glioblastomas. BMC Cell Biol 6: 7. doi: 10.1186/1471-2121-6-7
|
| [56] |
Demuth T, Berens ME (2004) Molecular mechanisms of glioma cell migration and invasion. J Neurooncol 70: 217–228. doi: 10.1007/s11060-004-2751-6
|
| [57] | Knott JC, Mahesparan R, Garcia-Cabrera I, et al. (1998) Stimulation of extracellular matrix components in the normal brain by invading glioma cells. Int J cancer 75: 864–872. |
| [58] |
Giese A, Kluwe L, Laube B, et al. (1996) Migration of human glioma cells on myelin. Neurosurgery 38: 755–764. doi: 10.1227/00006123-199604000-00026
|
| [59] | Soroceanu L, Manning TJ, Sontheimer H (1999) Modulation of glioma cell migration and invasion using Cl- and K+ ion channel blockers. J Neurosci 19: 5942–5954. |
| [60] |
Merzak A, Pilkington GJ (1997) Molecular and cellular pathology of intrinsic brain tumours. Cancer Metastasis Rev 16: 155–177. doi: 10.1023/A:1005760726850
|
| [61] |
Gaspar LE, Fisher BJ, Macdonald DR, et al. (1992) Supratentorial malignant glioma: patterns of recurrence and implications for external beam local treatment. Int J Radiat Oncol Biol Phys 24: 55–57. doi: 10.1016/0360-3016(92)91021-E
|
| [62] |
Puchner MJ, Herrmann HD, Berger J, et al. (2000) Surgery, tamoxifen, carboplatin, and radiotherapy in the treatment of newly diagnosed glioblastoma patients. J Neurooncol 49: 147–155. doi: 10.1023/A:1026533016912
|
| [63] |
Giese A, Bjerkvig R, Berens ME, et al. (2003) Cost of migration: invasion of malignant gliomas and implications for treatment. J Clin Oncol 21: 1624–1636. doi: 10.1200/JCO.2003.05.063
|
| [64] |
Sanai N, Polley M-Y, McDermott MW, et al. (2011) An extent of resection threshold for newly diagnosed glioblastomas. J Neurosurg 115: 3–8. doi: 10.3171/2011.2.JNS10998
|
| [65] |
Duffau H (2014) Diffuse low-grade gliomas and neuroplasticity. Diagn Interv Imaging 95: 945–955. doi: 10.1016/j.diii.2014.08.001
|
| [66] | Duffau H (2015) Resecting diffuse low-grade gliomas to the boundaries of brain functions: a new concept in surgical neuro-oncology. J Neurosurg Sci 59: 361–371. |
| [67] |
McGirt MJ, Mukherjee D, Chaichana KL, et al. (2009) Association of surgically acquired motor and language deficits on overall survival after resection of glioblastoma multiforme. Neurosurgery 65: 463–469. doi: 10.1227/01.NEU.0000349763.42238.E9
|
| [68] |
Hauser SB, Kockro RA, Actor B, et al. (2016) Combining 5-aminolevulinic acid fluorescence and intraoperative magnetic resonance imaging in glioblastoma surgery: A histology-based evaluation. Neurosurgery 78: 475–483. doi: 10.1227/NEU.0000000000001035
|
| [69] | Jaber M, Wölfer J, Ewelt C, et al. (2015) The Value of 5-ALA in Low-grade Gliomas and High-grade Gliomas Lacking Glioblastoma Imaging Features. Neurosurgery 78: 401–411. |
| [70] |
Stummer W, Pichlmeier U, Meinel T, et al. (2006) Fluorescence-guided surgery with 5-aminolevulinic acid for resection of malignant glioma: a randomised controlled multicentre phase III trial. Lancet Oncol 7: 392–401. doi: 10.1016/S1470-2045(06)70665-9
|
| [71] |
Sanai N, Berger MS (2008) G Lioma E Xtent of R Esection and and Methods. Neurosurgery 62: 753–766. doi: 10.1227/01.neu.0000318159.21731.cf
|
| [72] |
De Benedictis A, Moritz-Gasser S, Duffau H (2010) Awake mapping optimizes the extent of resection for low-grade gliomas in eloquent areas. Neurosurgery 66: 1074–1084. doi: 10.1227/01.NEU.0000369514.74284.78
|
| [73] | Desmurget M, Bonnetblanc F, Duffau H (2007) Contrasting acute and slow-growing lesions: a new door to brain plasticity. Brain 130: 898–914. |
| [74] |
Pallud J, Audureau E, Blonski M, et al. (2014) Epileptic seizures in diffuse low-grade gliomas in adults. Brain 137: 449–462. doi: 10.1093/brain/awt345
|
| [75] |
De Witt Hamer PC, Robles SG, Zwinderman AH, et al. (2012) Impact of intraoperative stimulation brain mapping on glioma surgery outcome: a meta-analysis. J Clin Oncol 30: 2559–2565. doi: 10.1200/JCO.2011.38.4818
|
| [76] |
Chang EF, Clark A, Smith JS, et al. (2011) Functional mapping-guided resection of low-grade gliomas in eloquent areas of the brain: improvement of long-term survival. Clinical article. J Neurosurg 114: 566–573. doi: 10.3171/2010.6.JNS091246
|
| [77] |
Sanai N, Polley M-Y, Berger MS (2010) Insular glioma resection: assessment of patient morbidity, survival, and tumor progression. J Neurosurg 112: 1–9. doi: 10.3171/2009.6.JNS0952
|
| [78] |
Lee WA, Bonin V, Reed M, et al. (2016) Anatomy and function of an excitatory network in the visual cortex. Nature 532: 1–18. doi: 10.1038/532S1a
|
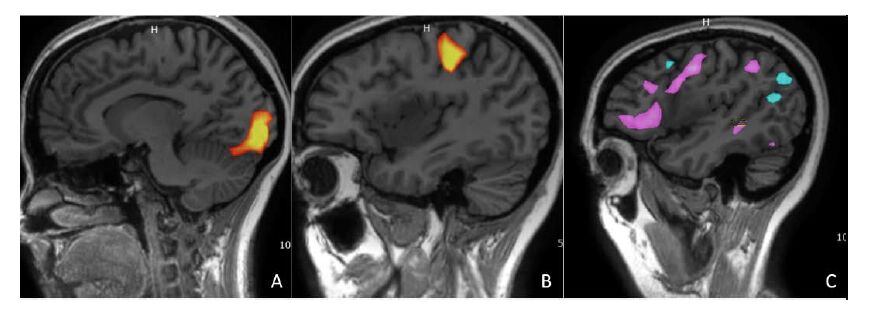
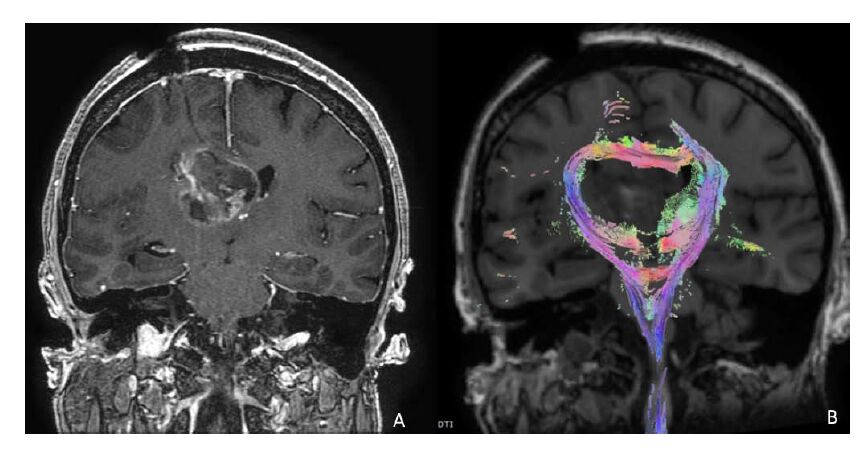
Figures(2)

Sanjay Konakondla, Steven A. Toms. Cerebral Connectivity and High-grade Gliomas: Evolving Concepts of Eloquent Brain in Surgery for Glioma[J]. AIMS Medical Science, 2017, 4(1): 52-70. doi: 10.3934/medsci.2017.1.52







 DownLoad:
DownLoad: