Citation: Karolína M. Šišková, Renáta Večeřová, Hana Kubičková, Magdaléna Bryksová, Klára Čépe, Milan Kolář. Revisiting spontaneous silver nanoparticles formation: a factor influencing the determination of minimum inhibitory concentration values?[J]. AIMS Environmental Science, 2015, 2(3): 607-622. doi: 10.3934/environsci.2015.3.607

| [1] |
Liu HH., Cohen Y (2014) Multimedia environmental distribution of engineered nanomaterials. Environ Sci Technol 48: 3281-3292. doi: 10.1021/es405132z

|
| [2] | Lopez-Serrano A, Olivas RM, Landaluze JS, et al. (2014) Nanoparticles: a global vision. Characterization, separation, and quantification methods. Potential environmental and health impact. Anal Methods 6: 38-56. |
| [3] |
Li DW, Zhai WL, Li YT, Long YT (2014) Recent progress in surface enhanced Raman spectroscopy for the detection of environmental pollutants. Microchim Acta 181: 23-43. doi: 10.1007/s00604-013-1115-3
|
| [4] |
Lalley J, Dionysiou DD, Varma RS, et al. (2014) Silver-based antibacterial surfaces for drinking water disinfection - an overview. Curr Opin Chem Eng 3: 25-29. doi: 10.1016/j.coche.2013.09.004
|
| [5] |
Han Ch, Likodimos V, Khan JA, et al. (2014) UV-visible Light-activated Ag-decorated, monodispersed TiO2 aggregates for treatment of the pharmaceutical oxytetracycline. Environ Sci Pollut Res 21: 11781-11793. doi: 10.1007/s11356-013-2233-5
|
| [6] |
Windler I, Height M, Nowack B (2013) Comparative evaluation of antimicrobials for textile applications. Environ Int 53: 62-73. doi: 10.1016/j.envint.2012.12.010
|
| [7] |
Geranio L, Heuberger M, Nowack B (2009) The behavior of silver nanotextiles during washing. Environ Sci Technol 43: 8113-8. doi: 10.1021/es9018332
|
| [8] |
Adegboyega NF, Sharma VK, Siskova K, et al. (2013) Interactions of Aqueous Ag+ with Fulvic Acids: Mechanisms of Silver Nanoparticle Formation and Investigation of Stability. Environ Sci Technol 47: 757-764. doi: 10.1021/es302305f
|
| [9] |
Akaighe N, MacCuspie RI, Navarro DA, et al. (2011) Humic acid-induced silver nanoparticle formation under environmentally relevant conditions. Environ Sci Technol 45: 3895-3901. doi: 10.1021/es103946g
|
| [10] |
Nadagouda MN, Iyanna N, Lalley J, et al. (2014) Synthesis of silver and gold nanoparticles using antioxidants from blackberry, blueberry, pomegranate, and turmeric extracts. ACS Sustanaible Chem Eng 2: 1717-1723. doi: 10.1021/sc500237k
|
| [11] |
Markova Z, Siskova KM, Filip J, et al. (2013) Air stable magnetic bimetallic Fe-Ag nanoparticles for advanced antimicrobial treatment and phosphorus removal. Environ Sci Technol 47: 5285-5293. doi: 10.1021/es304693g
|
| [12] | Sigma-Aldrich. Inc., Product Information, 70192 Mueller Hinton Broth (M-H Broth). Available from: http://www.sigmaaldrich.com/content/dam/sigma-aldrich/docs/Fluka/Datasheet/70192dat.pdf |
| [13] |
Sintubin L, Verstraete W, Boon N (2012) Biologically produced nanosilver: current state and future perspectives. Biotechnol Bioeng 109: 2422-2436. doi: 10.1002/bit.24570
|
| [14] |
Faramarzi MA, Sadighi A (2013) Insights into biogenic and chemical production of inorganic nanomaterials and nanostructures. Adv Colloid Interface Sci 189-190: 1-20. doi: 10.1016/j.cis.2012.12.001
|
| [15] |
Suman TY, Radhika Rajasree SR, Kanchana A, et al. (2013) Biosynthesis, characterization and cytotoxic effect of plant mediated silver nanoparticles using Morinda citrifolia root extract. Colloids Surf B 106: 74-78. doi: 10.1016/j.colsurfb.2013.01.037
|
| [16] |
Kharissova OV, Dias HVR, Kharisov BI, et al. (2013) The greener synthesis of nanoparticles. Trends Biotechnol 31: 240-248. doi: 10.1016/j.tibtech.2013.01.003
|
| [17] |
Hu B, Wang SB, Wang K, et al. (2008) Microwave-assisted rapid facile “green” synthesis of uniform silver nanoparticles: self-assembly into multilayered films and their optical properties. J Phys Chem C 112: 11169-11174. doi: 10.1021/jp801267j
|
| [18] |
Alvarez-Puebla RA, Aroca RF (2009) Synthesis of silver nanoparticles with controllable surface charge and their application to surface-enhanced Raman scattering. Anal Chem 81: 2280-2295. doi: 10.1021/ac8024416
|
| [19] |
Khan Z, Talib A (2010) Growth of different morphologies (quantum dots to nanorod) of Ag-nanoparticles: role of cysteine concentration. Coll Surf B 76: 164-169. doi: 10.1016/j.colsurfb.2009.10.029
|
| [20] |
Rafey A, Shrivastavaa KBL, Iqbal SA, et al. (2011) Growth of Ag-nanoparticles using aspartic acid in aqueous solutions. J Colloid Interface Sci 354: 190-195. doi: 10.1016/j.jcis.2010.10.046
|
| [21] |
Jacob JA, Naumov S, Mukherjee T, et al. (2011) Preparation, characterization, surface modification and redox reactions of silver nanoparticles in the presence of tryptophan. Coll Surf B 87: 498-504. doi: 10.1016/j.colsurfb.2011.06.017
|
| [22] |
Sondi I, Goia DV, Matijevic E (2003) Preparation of highly concentrated stable dispersions of uniform silver nanoparticles. J Colloid Interface Sci 260: 75-81. doi: 10.1016/S0021-9797(02)00205-9
|
| [23] | Nadagouda MN, Varma RS (2008) Green synthesis of Ag and Pd nanospheres, nanowires, and nanorods using vitamin B2: catalytic polymerisation of aniline and pyrrole. J Nanomat 2008: 1-8. |
| [24] |
Tan S, Erol M, Attygalle A, et al. (2007) Synthesis of positively charged silver nanoparticles via photoreduction of AgNO3 in branched polyethyleneimine/HEPES solutions. Langmuir 23: 9836-9843. doi: 10.1021/la701236v
|
| [25] |
Frattini A, Pellegri N, Nicastro D, et al. (2005) Effect of amine groups in the synthesis of Ag nanoparticles using aminosilanes. Mater Chem Phys 94: 148-152. doi: 10.1016/j.matchemphys.2005.04.023
|
| [26] |
Xie J, Lee JY, Wang DIC, et al. (2007) Silver nanoplates: from biological to biomimetic synthesis. ACS Nano 1: 429-439. doi: 10.1021/nn7000883
|
| [27] | Stevanovic M, Savanovic I, Uskokovic V, et al. (2012) A new, simple, green, and one-pot four-component synthesis of bare and poly(alpha,gamma, L-glutamic acid)-capped silver nanoparticles. Colloid Polym Sci 190: 221-231. |
| [28] |
Yu DG (2007) Formation of colloidal silver nanoparticles stabilized by Na+-poly(gamma-glutamic acid)-silver nitrate complex via chemical reduction process. Coll Surf B 59: 171-178. doi: 10.1016/j.colsurfb.2007.05.007
|
| [29] |
Vigneshwaran N, Nachane RP, Balasubramanya RH (2006) A novel one-pot “green” synthesis of stable silver nanoparticles using soluble starch. Carbohydr Res 341: 2012-2018. doi: 10.1016/j.carres.2006.04.042
|
| [30] |
Raveendran P, Fu J, Wallen SL (2003) Completely “green” synthesis and stabilization of metal nanoparticles. J Am Chem Soc 125: 13940-13941. doi: 10.1021/ja029267j
|
| [31] |
Tai C, Wang YH, Liu HS (2008) A green process for preparing silver nanoparticles using spinning disk reactor. AIChE J 54: 445-452. doi: 10.1002/aic.11396
|
| [32] |
Sun SK, Wang HF, Yan XP (2011) A sensitive and selective resonance light scattering bioassay for homocysteine in biological fluids based on target-involved assembly of polyethyleneimine-capped Ag-nanoclusters. Chem Commun 47: 3817-3819. doi: 10.1039/c0cc04463f
|
| [33] |
Lee KJ, Browning LM, Nallathamby PD, et al. (2013) Study of charge-dependent transport and toxicity of peptide-functionalized silver nanoparticles using zebrafish embryos and single nanoparticle plasmonic spectroscopy. Chem Res Toxicol 26: 904-917. doi: 10.1021/tx400087d
|
| [34] |
Chernousova S, Epple M (2013) Silver as antibacterial agent: ion, nanoparticle, and metal. Angew Chem Ind Ed 52: 1636-1653. doi: 10.1002/anie.201205923
|
| [35] |
Sharma VK, Siskova KM, Zboril R, et al. (2014) Organic-coated silver nanoparticles in biological and environmental conditions: Fate, stability and toxicity. Adv Colloid Interface Sci 204: 15-34. doi: 10.1016/j.cis.2013.12.002
|
| [36] |
Rai MK, Deshmukh SD, Ingle AP, et al. (2012) Silver nanoparticles: the powerful nanoweapon against multidrug-resistant bacteria. J Appl Microbiol 112: 841-852. doi: 10.1111/j.1365-2672.2012.05253.x
|
| [37] |
Li WR, Xie XB, Shi QS, et al. (2010) Antibacterial activity and mechanism of silver nanoparticles on Escherichia coli. Appl Microbiol Biotechnol 85: 1115-1122. doi: 10.1007/s00253-009-2159-5
|
| [38] |
Li WR, Xie XB, Shi QS, et al. (2011) Antibacterial effect of silver nanoparticles on Staphylococcus aureus. Biometals 24: 135-141. doi: 10.1007/s10534-010-9381-6
|
| [39] |
Lara HH, Ayala-Nunez NV, Turrent LCI, et al. (2010) Bactericidal effect of silver nanoparticles against multidrug-resistant bacteria. World J Microbiol Biotechnol 26: 615-621. doi: 10.1007/s11274-009-0211-3
|
| [40] |
Lok CN, Ho CM, Chen R, et. al. (2006) Proteomic analysis of the mode of antibacterial action of silver nanoparticles. J Proteome Res 5: 916-924. doi: 10.1021/pr0504079
|
| [41] |
Yang XY, Gondikas AP, Marinakos SM, et al. (2012) Mechanism of silver nanoparticle toxicity is dependent on dissolved silver and surface coating in Caenorhabditis elegans. Environ Sci Technol 46: 1119-1127. doi: 10.1021/es202417t
|
| [42] |
Morones JR, Elechiguerra, JL, Camacho A, et al. (2005) The bactericidal effect of silver nanoparticles. Nanotechnology 16: 2346. doi: 10.1088/0957-4484/16/10/059
|
| [43] | Sondi I, Salopek-Sondi B (2004) Silver nanoparticles as antimicrobial agent: a case study on E. coli as a model for gram-negative bacteria. J Colloid Interface Sci 275: 177-182. |
| [44] |
Xu H, Qu F, Xu H, et al. (2012) Role of reactive oxygen species in the antibacterial mechanism of silver nanoparticles on Escherichia coli O157:H7. Biometals 25: 45-53. doi: 10.1007/s10534-011-9482-x
|
| [45] |
Choi O, Hu Z (2008) Size dependent and reactive oxygen species related nanosilver toxicity to nitrifying bacteria. Environ Sci Technol 42: 4583-4588. doi: 10.1021/es703238h
|
| [46] |
Xiu ZM, Zhang QB, Puppala HL, et al. (2012) Negligible particle-specific antibacterial activity of silver nanoparticles. Nano Lett 12: 4271-4275. doi: 10.1021/nl301934w
|
| [47] |
El Badawy AM, Luxton TP, Silva RG, et al. (2010) Impact of environmental conditions (pH, ionic strength, and electrolyte type) on the surface charge and aggregation of silver nanoparticles suspensions. Environ Sci Technol 44: 1260-1266. doi: 10.1021/es902240k
|
| [48] |
Siskova KM, Machala L, Tucek J, et al. (2013) Mixtures of L-amino acids as reaction medium for iron nanoparticles formation: the order of addition into ferrous salt solution matters. Int J Mol Sci 14: 19452-19473. doi: 10.3390/ijms141019452
|
| [49] |
Sloufova I, Siskova K, Vlckova B, et al. (2008) SERS-activating effect of chlorides on borate-stabilized silver nanoparticles: formation of new reduced adsorption sites and induced nanoparticle fusion. Phys Chem Chem Phys 10: 2233-2242. doi: 10.1039/b718178g
|
| [50] |
Siskova K, Becicka O, Safarova K, et al. (2013) HCl effect on two types of Ag nanoparticles utilizable in detection of low concentrations of organic species. Sustainable nanotechnology and the environment: advances and achievements 1124: 151-163. doi: 10.1021/bk-2013-1124.ch009
|
| [51] |
Sriramulu DD, Lunsdorf H, Lam JS, et al. (2005) Microcolony formation: a novel biofilm model of Pseudomonas aeruginosa for the cystic fibrosis lung. J Med Microbiol 54: 667-676. doi: 10.1099/jmm.0.45969-0
|
| [52] | Nakamoto K (2009) Infrared and Raman spectra of inorganic and coordination compounds, part B, Hoboken, New Jersey, USA, John Wiley & Sons, Inc., 26-84 and 388-392. |
| [53] |
Moskovits M, Suh JS (1985) Confirmation of mono- and dicarboxylic acids adsorbed on silver surfaces. J Am Chem Soc 107: 6826-6829. doi: 10.1021/ja00310a014
|
| [54] |
Munro CH, Smith WE, Garner M, et al. (1995) Characterization of the surface of a citrate-reduced colloid optimized for use as a substrate for surface-enhanced resonance Raman scattering. Langmuir 11: 3712-3720. doi: 10.1021/la00010a021
|
| [55] | Lindon JC (2000) Encyclopedia of spectroscopy and spectrometry, part II, San Diego, Academic press, 1035-1057. |
| [56] | Schaumann GE, Philippe A, Bundschuh M, et al. (2014) Understanding the fate and biological effects of Ag- and TiO2- nanoparticles in the environment: The quest for advanced analytics and interdisciplinary concepts. Sci Total Environ: S0048-9697: 1473-1479. |
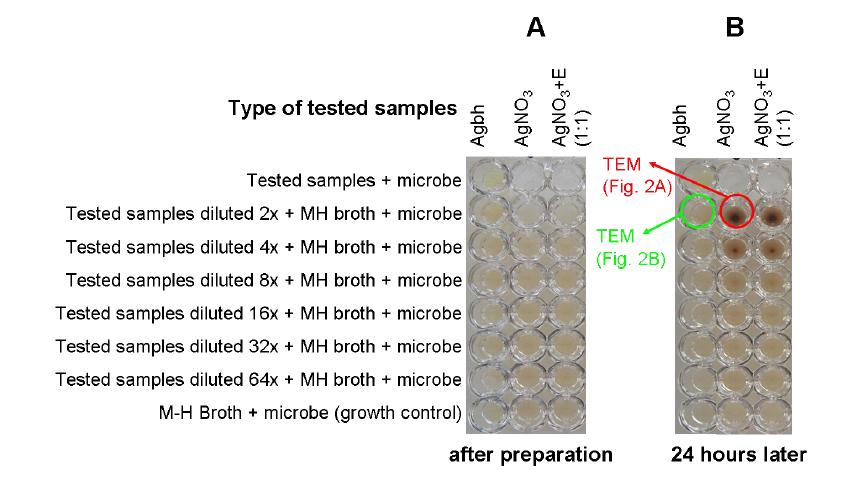
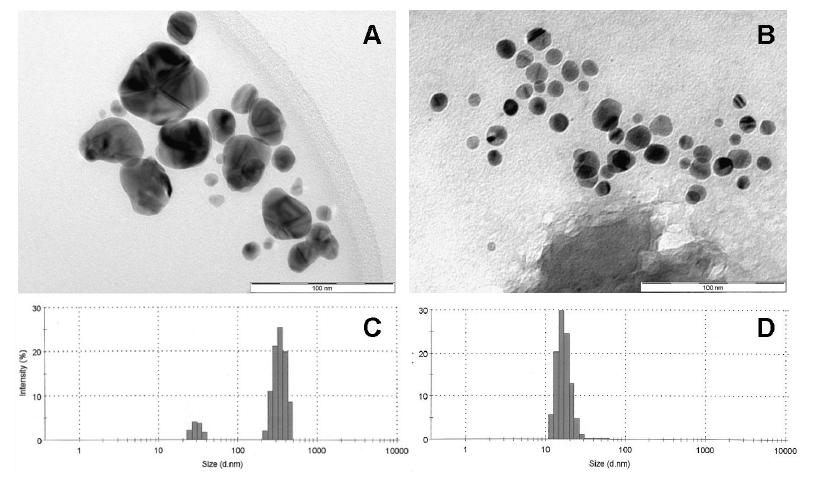
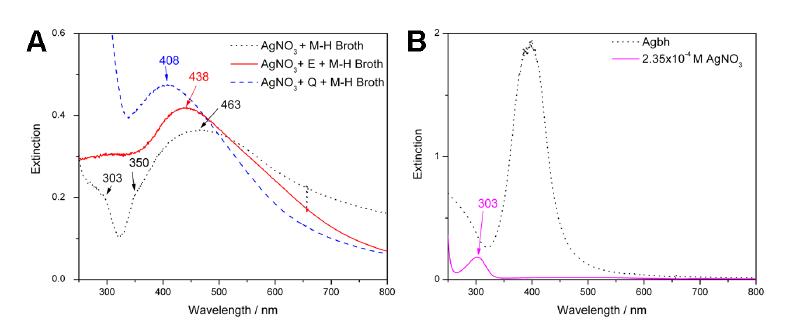
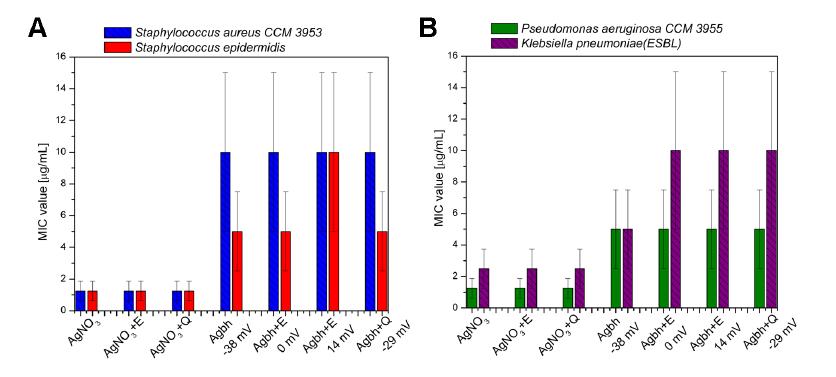
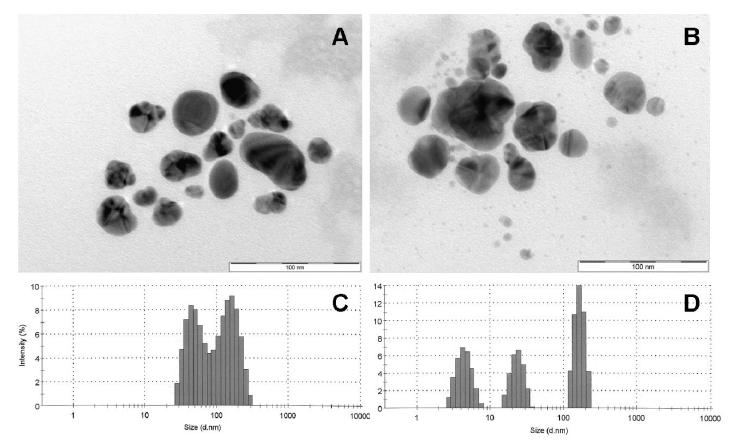
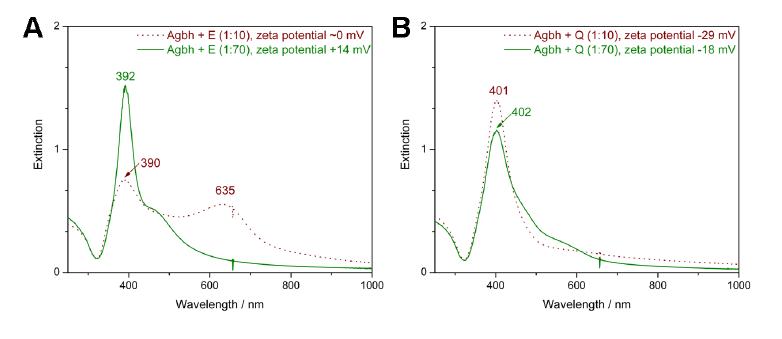
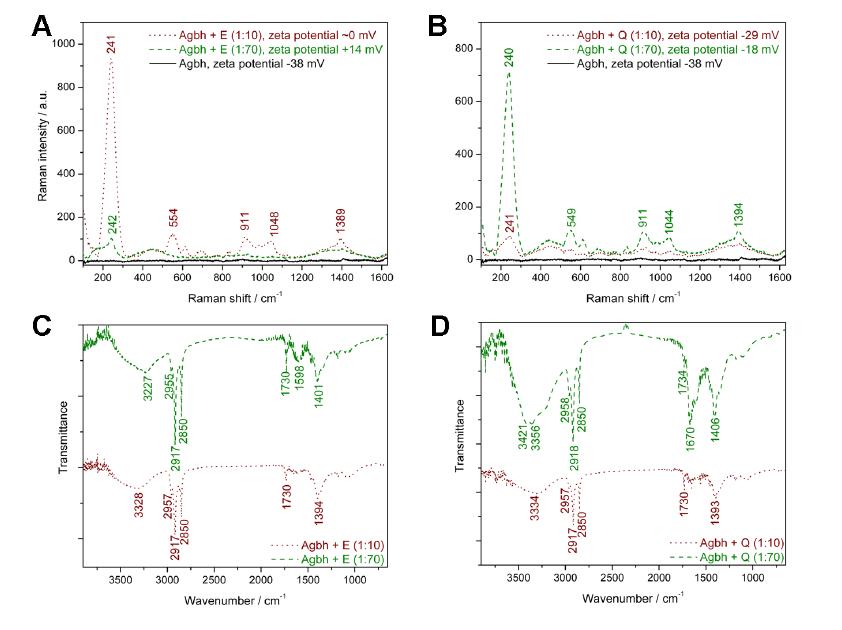
Figures(7)

Karolína M. Šišková, Renáta Večeřová, Hana Kubičková, Magdaléna Bryksová, Klára Čépe, Milan Kolář. Revisiting spontaneous silver nanoparticles formation: a factor influencing the determination of minimum inhibitory concentration values?[J]. AIMS Environmental Science, 2015, 2(3): 607-622. doi: 10.3934/environsci.2015.3.607







 DownLoad:
DownLoad: