Citation: Amira Bouhali, Walid Ben Aribi, Slimane Ben Miled, Amira Kebir. Impact of immunity loss on the optimal vaccination strategy for an age-structured epidemiological model[J]. Mathematical Biosciences and Engineering, 2024, 21(6): 6372-6392. doi: 10.3934/mbe.2024278

| [1] |
N. Wu, K. Joyal-Desmarais, P. A. B. Ribeiro, A. M. Vieira, J. Stojanovic, C. Sanuade, et al., Long-term effectiveness of COVID-19 vaccines against infections, hospitalizations, and mortality in adults: findings from a rapid living systematic evidence synthesis and meta-analysis up to December 2022, Lancet Respir. Med., 11 (2023), 439–452. https://doi.org/10.1016/S2213-2600(23)00015-2 doi: 10.1016/S2213-2600(23)00015-2

|
| [2] |
A. I. Abioye, O. J. Peter, H. A. Ogunseye, F. A. Oguntolu, K. Oshinubi, A. A. Ibrahim, et al., Mathematical model of COVID-19 in Nigeria with optimal control, Res. Phys., 28 (2021), 104598. https://doi.org/10.1016/j.rinp.2021.104598 doi: 10.1016/j.rinp.2021.104598
|
| [3] |
M. Diarra, A. Kebir, C. Talla, A. Barry, J. Faye, D. Louati, et al., Non-pharmaceutical interventions and COVID-19 vaccination strategies in Senegal: a modelling study, BMJ Glob. Health, 7 (2022), e007236. https://doi.org/10.1136/bmjgh-2021-007236 doi: 10.1136/bmjgh-2021-007236
|
| [4] |
G. Gonzalez-Parra, M. R. Cogollo, A. J. Arenas, Mathematical modeling to study optimal allocation of vaccines against COVID-19 using an age-structured population, Axioms, 11 (2022), 109. https://doi.org/10.3390/axioms11030109 doi: 10.3390/axioms11030109
|
| [5] |
A. B. Hogan, P. Winskill, O. J. Watson, P. G. T. Walker, C. Whittaker, M. Baguelin, et al., Within-country age-based prioritisation, global allocation, and public health impact of a vaccine against SARS-CoV-2: A mathematical modelling analysis, Vaccine, 39 (2021), 2995–3006. https://doi.org/10.1016/j.vaccine.2021.04.002 doi: 10.1016/j.vaccine.2021.04.002
|
| [6] |
P. Jia, J. Yang, X. Li, Optimal control and cost-effective analysis of an age-structured emerging infectious disease model, Inf. Dis. Modell., 1 (2021), 149–169. https://doi.org/10.1016/j.idm.2021.12.004 doi: 10.1016/j.idm.2021.12.004
|
| [7] | A. A. Khan, S. Ullah, R. Amin, Optimal control analysis of COVID-19 vaccine epidemic model: a case study, Eur. Phys. J. Plus, 137 (2022). https://doi.org/10.1140/epjp/s13360-022-02365-8 |
| [8] |
L. Matrajt, J. Eaton, T. Leung, D. Dimitrov, J. T. Schiffer, D. A. Swan, et al., optimising vaccine allocation for COVID-19 vaccines shows the potential role of single-dose vaccination, Nat. Commun., 12 (2021), 3449. https://doi.org/10.1038/s41467-021-23761-1 doi: 10.1038/s41467-021-23761-1
|
| [9] |
M. M. Ojo, T. O. Benson, O. J. Peter, E. F. Doungmo Goufo, Nonlinear optimal control strategies for a mathematical model of COVID-19 and influenza co-infection, Phys. A: Stat. Mech. Appl., 607 (2022), 128173. https://doi.org/10.1016/j.physa.2022.128173 doi: 10.1016/j.physa.2022.128173
|
| [10] |
A. Olivares, E. Staffetti, Optimal control-based vaccination and testing strategies for COVID-19, Comput. Methods Programs Biomed., 211 (2021), 106411. https://doi.org/10.1016/j.cmpb.2021.106411 doi: 10.1016/j.cmpb.2021.106411
|
| [11] |
M. Urban, J. Jodlowska, J. Balbus, K. Kubica, Vaccination strategies based on a mathematical model of epidemics considering the age structure of the population, WSEAS Trans. Biol Biomed., 21 (2024), 29–39. https://doi.org/10.37394/23208.2024.21.4 doi: 10.37394/23208.2024.21.4
|
| [12] |
S. Zhou, S. Zhou, Z. Zheng, J. Lu, optimising spatial allocation of COVID-19 vaccine by agent-based spatiotemporal simulations, Geohealth, 6 (2021), e2021GH000427. https://doi.org/10.1029/2021GH000427 doi: 10.1029/2021GH000427
|
| [13] | O. Diekmann J. A. P. Heesterbeek, Mathematical Epidemiology of Infectious Diseases: Model Building, Analysis and Interpretation, John Wiley & Sons, 2000. |
| [14] |
T. Britton, F. Ball, P. Trapman, A mathematical model reveals the influence of population heterogeneity on herd immunity to SARS-CoV-2, Science, 369 (2020), 846–849. https://doi.org/10.1126/science.abc6810 doi: 10.1126/science.abc6810
|
| [15] |
S. Bentout, A. Tridane, S. Djilali, T. M. Touaoula, Age-structured modeling of COVID-19 epidemic in the USA, UAE and Algeria, Alex. Eng. J., 60 (2021), 401–411. https://doi.org/10.1016/j.aej.2020.08.053 doi: 10.1016/j.aej.2020.08.053
|
| [16] |
J. Read, J. Lessler, S. Riley, S. Wang, L. Tan, K. Kwok, et al., Social mixing patterns in rural and urban areas of southern China, Proc. Biol. Sci., 281 (2014), 122. https://doi.org/10.1098/rspb.2014.0268 doi: 10.1098/rspb.2014.0268
|
| [17] |
I. Voinsky, G. Baristaite, D. Gurwitz, Effects of age and sex on recovery from COVID-19: Analysis of 5769 Israeli patients, J. Infect., 2 (2020), e102–e103. https://doi.org/10.1016/j.jinf.2020.05.026 doi: 10.1016/j.jinf.2020.05.026
|
| [18] |
S. S. Shen-Orr, D. Furman, Variability in the immune system: of vaccine responses and immune states, Curr. Opin. Immunol., 4 (2013), 542–547. https://doi.org/10.1016/j.coi.2013.07.009 doi: 10.1016/j.coi.2013.07.009
|
| [19] |
L. L. Dietz, A. K. Juhl, O. S. Søgaard, J. Reekie, H. Nielsen, I. S. Johansen, et al., Impact of age and comorbidities on SARS-CoV-2 vaccine-induced T cell immunity, Commun. Med., 1 (2023), 58. https://doi.org/10.1038/s43856-023-00277-x doi: 10.1038/s43856-023-00277-x
|
| [20] |
B. Naffeti, W. Ben Aribi, A. Kebir, M. Diarra, M. Schoenhals, I. Vigan-Womas, et al., Comparative reconstruction of SARS-CoV-2 transmission in three African countries using a mathematical model integrating immunity data, IJID Reg., 10 (2024), 100–107. https://doi.org/10.1016/j.ijregi.2023.11.011 doi: 10.1016/j.ijregi.2023.11.011
|
| [21] | A. Bouhali, W. Ben Aribi, A. Kebir, S. Ben Miled, Age optimal vaccination strategy for respiratory infectious disease: a constraint-dependant approach, Authorea Prepr., 2023. https://doi.org/10.22541/au.169571176.62627085/v1 |
| [22] |
E. H. Elbasha, A. B. Gumel, Vaccination and herd immunity thresholds in heterogeneous populations, J. Math. Biol., 83 (2021), 73. https://doi.org/10.1007/s00285-021-01686-z doi: 10.1007/s00285-021-01686-z
|
| [23] |
K. Hattaf, M. I. El Karimi, A. A. Mohsen, Z. Hajhouji, M. El Younoussi, N. Yousfi, Mathematical modeling and analysis of the dynamics of RNA viruses in presence of immunity and treatment: A case study of SARS-CoV-2, Vaccines, 11 (2023), 2076–393X. https://doi.org/10.3390/vaccines11020201 doi: 10.3390/vaccines11020201
|
| [24] |
B. Naffeti, S. Bourdin, W. Ben Aribi, A. Kebir, S. Ben Miled, Spatio-temporal evolution of the COVID-19 across African countries, Front. Public Health, 28 (2022), 1039925. https://doi.org/10.3389/fpubh.2022.1039925 doi: 10.3389/fpubh.2022.1039925
|
| [25] |
N. G. Davies, P. Klepac, Y. Liu, K. Prem, M. Jit, R. M. Eggo, Age-dependent effects in the transmission and control of COVID-19 epidemics, Nat. Med., 26 (2020), 1205–1211. https://doi.org/10.1038/s41591-020-0962-9 doi: 10.1038/s41591-020-0962-9
|
| [26] | WHO, Global-covid-19-vaccination-mid-2022, 2022. Available from: https://reliefweb.int/report/world/strategy-achieve-global-covid-19-vaccination-mid-2022. |
| [27] | W. Fleming, R. Rishel, Deterministic and Stochastic Optimal Control, Springer-Verlag, New York Heidelberg Berlin, 1986. |
| [28] | E. Trelat, Contrôle Optimal : Théorie et Applications, Université Pierre et Marie Curie (Paris 6) et Institut Universitaire de France, Paris, 2005. |
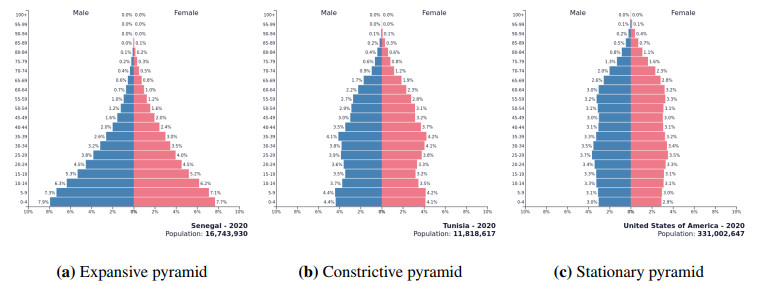
| [29] | Population Pyramids of the World from 1950 to 2100, Population Pyramid.net, 2020. Available from: https://www.populationpyramid.net/tunisia/2020/. |
| [30] | Stratégie Vaccinale Contre la COVID-19 en Tunisie, Report of the Tunisian Ministry of Public Health for COVID-19 Vaccination, 2021. Available from: http://www.santetunisie.rns.tn/images/strategie-vaccination-covid-19.pdf. |
| [31] |
K. Prem, A. R. Cook, M. Jit, Projecting social contact matrices in 152 countries using contact surveys and demographic data, PLoS Comput. Biol., 13 (2017), e1005697. https://doi.org/10.1371/journal.pcbi.1005697 doi: 10.1371/journal.pcbi.1005697
|
| [32] | WHO, COVID-19 Weekly Epidemiological Update, Available from: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19—10-august-2023. |
| [33] | WHO, WHO COVID-19 dashboard, Available from: https://data.who.int/dashboards/covid19/cases?n = c. |
| [34] | Our World in Data, Number of Deaths by Age, World, 2021. Available from: https://ourworldindata.org/grapher/annual-deaths-by-age. |
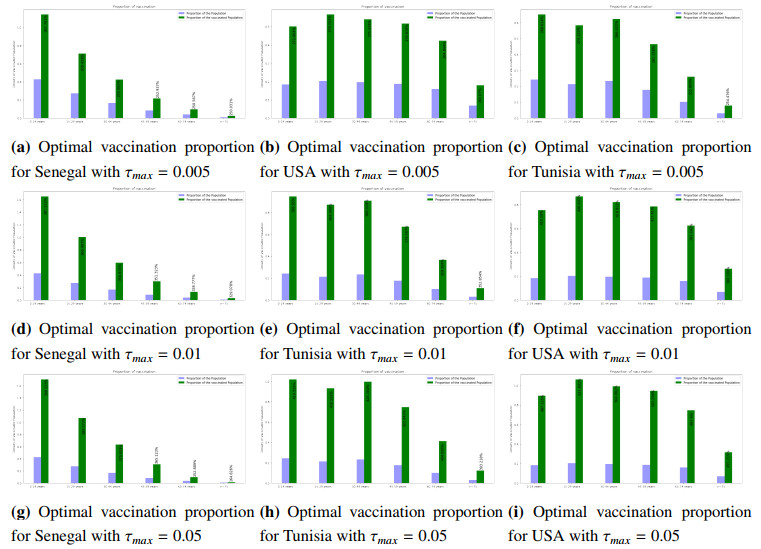
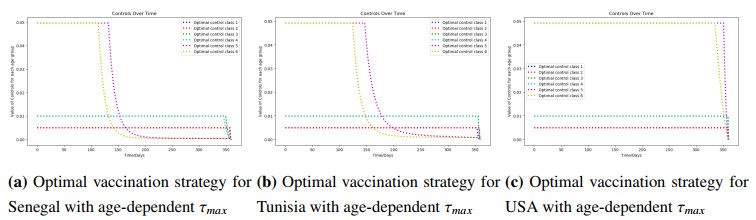
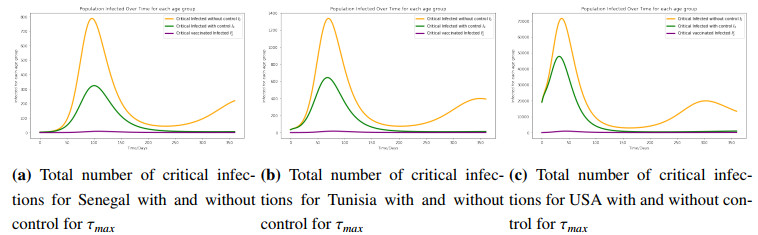
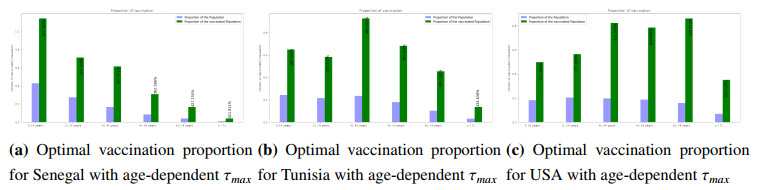
Figures(8) / Tables(3)

Amira Bouhali, Walid Ben Aribi, Slimane Ben Miled, Amira Kebir. Impact of immunity loss on the optimal vaccination strategy for an age-structured epidemiological model[J]. Mathematical Biosciences and Engineering, 2024, 21(6): 6372-6392. doi: 10.3934/mbe.2024278







 DownLoad:
DownLoad: