We present a new epidemic model highlighting the roles of the immunization time and concurrent use of different vaccines in a vaccination campaign. To this aim, we introduce new intra-compartmental dynamics, a procedure that can be extended to various other situations, as detailed through specific case studies considered herein, where the dynamics within compartments are present and influence the whole evolution.
Citation: Rinaldo M. Colombo, Francesca Marcellini, Elena Rossi. Vaccination strategies through intra—compartmental dynamics[J]. Networks and Heterogeneous Media, 2022, 17(3): 385-400. doi: 10.3934/nhm.2022012
We present a new epidemic model highlighting the roles of the immunization time and concurrent use of different vaccines in a vaccination campaign. To this aim, we introduce new intra-compartmental dynamics, a procedure that can be extended to various other situations, as detailed through specific case studies considered herein, where the dynamics within compartments are present and influence the whole evolution.

| [1] |
Mathematical modeling of immunity to malaria. Nonlinearity in biology and medicine. Math. Biosci. (1988) 90: 385-396. 
|
| [2] |
A multiscale model of virus pandemic: Heterogeneous interactive entities in a globally connected world. Math. Models Methods Appl. Sci. (2020) 30: 1591-1651.
|
| [3] |
F. Brauer and C. Castillo-Chavez, Mathematical Models in Population Biology and Epidemiology, 2$^{nd}$ edition, Texts in Applied Mathematics, 40, Springer, New York, 2012. |
| [4] |
Models for transmission of disease with immigration of infectives. Math. Biosci. (2001) 171: 143-154.
|
| [5] |
Well posedness and control in a nonlocal SIR model. Appl. Math. Optim. (2021) 84: 737-771.
|
| [6] |
R. M. Colombo, M. Garavello, F. Marcellini and E. Rossi, An age and space structured SIR model describing the Covid-19 pandemic, J. Math. Ind., 10 (2020), 20pp. |
| [7] |
R. M. Colombo, M. Garavello, F. Marcellini and E. Rossi, IBVPs for inhomogeneous systems of balance laws in several space dimensions motivated by biology and epidemiology, preprint, 2021. |
| [8] |
G. Dimarco, L. Pareschi, G. Toscani and M. Zanella, Wealth distribution under the spread of infectious diseases, Phys. Rev. E, 102 (2020), 14pp. |
| [9] |
Bifurcation thresholds in an SIR model with information-dependent vaccination. Math. Model. Nat. Phenom. (2007) 2: 23-38.
|
| [10] |
S. Ghosh and S. Bhattacharya, A data-driven understanding of COVID-19 dynamics using sequential genetic algorithm based probabilistic cellular automata, Appl. Soft Comput., 96 (2020). |
| [11] |
Modelling the COVID-19 epidemic and implementation of population-wide interventions in Italy. Nature Medicine (2020) 26: 855-860.
|
| [12] |
A. Godio, F. Pace and A. Vergnano, SEIR modeling of the Italian epidemic of SARS-CoV-2 using computational swarm intelligence, Internat. J. Environ. Res. Public Health, 17 (2020). |
| [13] |
Some results for an SEIR epidemic model with density dependence in the death rate. IMA J. Math. Appl. Med. Biol. (1992) 9: 67-106.
|
| [14] |
H. Inaba, Age-structured SIR epidemic model, in Age-Structured Population Dynamics in Demography and Epidemiology, Springer, 2017,287–331. |
| [15] |
Contributions to the mathematical theory of epidemics. II. The problem of endemicity. Proc. Roy. Soc. Lond. A (1932) 138: 55-83.
|
| [16] |
A contribution to the mathematical theory of epidemics. Proc. Roy. Soc. Lond. A (1927) 115: 700-721.
|
| [17] |
A simple vaccination model with multiple endemic states. Math. Biosci. (2000) 164: 183-201.
|
| [18] |
Global stability of a SEIR epidemic model with infectious force in latent, infected and immune period. Chaos Solitons Fractals (2005) 25: 1177-1184.
|
| [19] |
Global dynamics an SEIR epidemic model with vertical transmission. SIAM J. Appl. Math. (2001) 62: 58-69.
|
| [20] |
SVIR epidemic models with vaccination strategies. J. Theoret. Biol. (2008) 253: 1-11.
|
| [21] |
Dynamic models of infectious diseases as regulators of population sizes. J. Math. Biol. (1992) 30: 693-716.
|
| [22] |
J. D. Murray, Mathematical Biology. I. An Introduction, 3$^{rd}$ edition, Interdisciplinary Applied Mathematics, 17, Springer-Verlag, New York, 2002. |
| [23] |
B. Perthame, Transport Equations in Biology, Frontiers in Mathematics, Birkhäuser Verlag, Basel, 2007. |
| [24] |
C. Piazzola, L. Tamellini and R. Tempone, A note on tools for prediction under uncertainty and identifiability of SIR-like dynamical systems for epidemiology, Math. Biosci., 332 (2021), 21pp. |
| [25] |
Epidemic and demographic interaction in the spread of potentially fatal diseases in growing populations. Math. Biosci. (1992) 111: 99-130.
|
| [26] |
H. Wackerhage, R. Everett, K. Krüger, M. Murgia and P. Simon, et al., Sport, exercise and COVID-19, the disease caused by the SARS-CoV-2 coronavirus, Dtsch. Z. Sportmed., 71 (2020), E1–E12. |
| [27] |
Using a genetic algorithm to fit parameters of a COVID-19 SEIR model for US states. Math. Comput. Simulation (2021) 185: 687-695.
|




Figures(5) / Tables(3)
Rinaldo M. Colombo, Francesca Marcellini, Elena Rossi. Vaccination strategies through intra—compartmental dynamics[J]. Networks and Heterogeneous Media, 2022, 17(3): 385-400. doi: 10.3934/nhm.2022012

Solutions to (3)–(7)–(8) in the
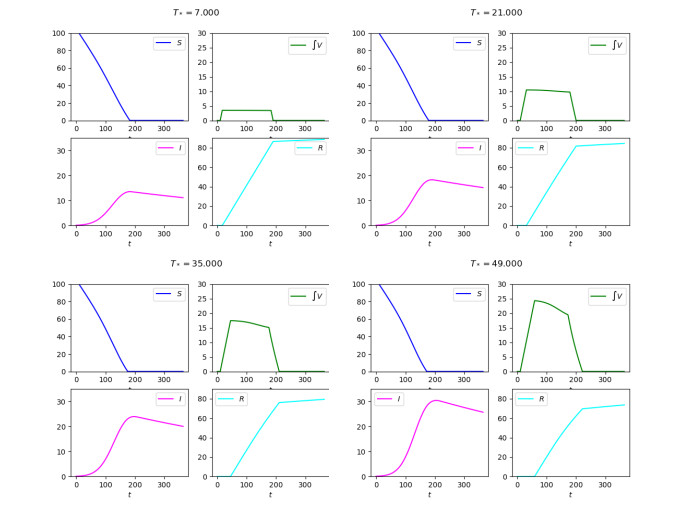
Diagrams of the solutions to (3)–(7)–(8) with a suspension in the vaccination campaign as detailed in (9) in the
Diagrams of the solutions to (4)–(7)–(8)–(10). On the left with
Above, the integrations of (1) and (15), below on the left that of (16) (19). The rightmost diagram on the second line displays the total number of living individuals in the three cases, showing that, with respect to mortality, the ODE–PDE model (16) can be seen in some senses in the middle between the ODE models (1) and (15)
Above, from left to right, the integrations of Case






 DownLoad:
DownLoad: