Citation: José Luis Alonso, Wolfgang H. Goldmann. Cellular mechanotransduction[J]. AIMS Biophysics, 2016, 3(1): 50-62. doi: 10.3934/biophy.2016.1.50

| [1] |
Wang JHC, Thampatty BP (2006) An introductory review in cell mechanobiology. Biomechan Model Mechanobiol 5: 1–6. doi: 10.1007/s10237-005-0012-z

|
| [2] | Osmanagic-Myers S, Dechat T, Foisner R (2015) Lamins at the crossroads of mechanosignaling. Genes Dev 29: 225–237. |
| [3] |
Bausch AR, Schwarz US (2013) Cellular mechanosensing: Sharing the force. Nat Mat 12: 948–949. doi: 10.1038/nmat3791
|
| [4] |
Luo T, Mohan K, Iglesias PA, et al. (2013) Molecular mechanisms of cellular mechanosensing. Nat Mat 12: 1064–1071. doi: 10.1038/nmat3772
|
| [5] | Wang N, Tytell JD, Ingber DE (2009) Mechanotransduction at a distance: mechanically coupling the extracellular matrix with the nucleus. Nat Rev Mol Cell Biology 10: 75–82. |
| [6] | Goldmann WH (2012a) Mechanotransduction in cells. Cell Biol Int 36: 649–652. |
| [7] | Shao X, Li Q, Mogilner A, et al. (2015) Mechanical stimulation induces formin-dependent assembly of a perinuclear actin rim. Proc Nat Acad Sci USA122: E2595–2601. |
| [8] |
Jalali S, del Pozo MA, Chen KD, et al. (2001) Integrin-mediated mechanotransduction requires its dynamic interaction with specific extracellular matrix (ECM) ligands. Proc Natl Acad Sci USA 98: 1042–1046. doi: 10.1073/pnas.98.3.1042
|
| [9] | Steinwachs J, Metzner C, Skodzek K, et al. (2015) Three-dimensional force microscopy of cells in biopolymer networks. Nat Methods [in press]. |
| [10] |
Geiger B, Spatz JP, Bershadsky AD (2009) Environmental sensing through focal adhesions. Nat Rev Molecular Cell Biology 10: 21–33. doi: 10.1038/nrm2593
|
| [11] |
Jaalouk DE, Lammerding J (2009) Mechanotransduction gone awry. Nat Rev Molecular Cell Biology 10: 63–73. doi: 10.1038/nrm2597
|
| [12] |
Grashoff C, Hofman BD, Brenner MD, et al. (2010) Measuring mechanical tension across vinculin reveals regulation of focal adhesion dynamics. Nature 466: 263–267. doi: 10.1038/nature09198
|
| [13] | Honarmandi P, Lee H, Lang MJ, et al. (2010) A microfluidic system with optical laser tweezers to study mechanotransduction and focal adhesion recruitment. Lab Chip 11: 684–694. |
| [14] |
Fabry B, Klemm AH, Kienle S, et al. (2011) Focal adhesion kinase stabilizes the cytoskeleton. Biophys J 101: 2131–2138. doi: 10.1016/j.bpj.2011.09.043
|
| [15] |
Goldmann WH (2014) Mechanosensation: a basic cellular process. Progress in Molecular Biology and Translational Science 126: 75–102. doi: 10.1016/B978-0-12-394624-9.00004-X
|
| [16] |
Dent JE, Devescovi V, Li H, et al. (2015) Mechanotransduction map: simulation model, molecular pathway, gene set. Bioinformatics 31: 1053–1059. doi: 10.1093/bioinformatics/btu776
|
| [17] | Goldmann WH (2016) Role of vinculin in cellular mechanotransduction. Cell Biol Int [in press]. |
| [18] |
Janoštiak R, Pataki AC, Brabek J, et al. (2014) Mechanosensors in integrin signaling: the emerging role of p130Cas. Eur J Cell Biol 93: 445–454. doi: 10.1016/j.ejcb.2014.07.002
|
| [19] |
Ezzell RM, Goldmann WH, Wang N, et al. (1997) Vinculin promotes cell spreading by mechanically coupling integrins to the cytoskeleton. Exp Cell Res 231: 14–26. doi: 10.1006/excr.1996.3451
|
| [20] |
Mierke CT, Kollmannsberger P, Zitterbart DP, et al. (2010) Vinculin facilitates cell invasion into three-dimensional collagen matrices. J Biol Chem 285: 13121–13130. doi: 10.1074/jbc.M109.087171
|
| [21] |
Wozniak MA, Chen CS (2009) Mechanotransduction in development: a growing role for contractility. Nat Rev Molecular Cell Biology 10: 34–42. doi: 10.1038/nrm2592
|
| [22] |
Bays JL, Peng X, Tolbert CE, et al. (2014) Vinculin phosphorylation differentially regulates mechanotransduction at cell-cell and cell-matrix adhesions. J Cell Biol 205: 251–263. doi: 10.1083/jcb.201309092
|
| [23] |
Martinac B (2004) Mechanosensitive ion channels: molecules of mechanotransduction. J Cell Sci 117: 2449–2460. doi: 10.1242/jcs.01232
|
| [24] |
Ingber DE (2006) Cellular mechanotransduction: putting all the pieces together again. FASEB J 20: 811–827. doi: 10.1096/fj.05-5424rev
|
| [25] | Haswell ES, Phillips R, Rees DC (2011) Mechanosensitive channels: what can they do and how do they do it? Structure 19: 1356–1369. |
| [26] |
Delmas P, Hao J, Rodat-Despoix L (2011) Molecular mechanisms of mechanotransduction in mammalian sensory neurons. Nat Rev Neurosci 12: 139–153. doi: 10.1038/nrn2993
|
| [27] |
Nomura S, Yamamoto TT (2000) Molecular events caused by mechanical stress in bone. Matrix Biology 19: 91–96. doi: 10.1016/S0945-053X(00)00050-0
|
| [28] |
Hoffman BD, Grashoff C, Schwartz MA (2011) Dynamic molecular processes mediate cellular mechanotransduction. Nature 475: 316–23. doi: 10.1038/nature10316
|
| [29] |
Luo T, Mohan K, Iglesias PA, et al. (2013) Molecular mechanisms of cellular mechanosensing. Nat Materials 12: 1064–1071. doi: 10.1038/nmat3772
|
| [30] |
Leerberg JM, Gomez GA, Verma S, et al. (2014) Tension-sensitive actin assembly supports contractility at the epithelial zonula adherens. Curr Biology 24: 1689–1699. doi: 10.1016/j.cub.2014.06.028
|
| [31] |
Milllward-Sadler SJ, Salter DM (2004) Integrin-dependent signal cascades in chondrocyte mechanotransduction. Ann Biomed Eng 32: 435–446. doi: 10.1023/B:ABME.0000017538.72511.48
|
| [32] |
Shivashankar GV (2011) Mechanosignaling to the cell nucleus and gene regulation. Ann Rev Biophysics 40: 361–378. doi: 10.1146/annurev-biophys-042910-155319
|
| [33] | McCain ML, Parker KK (2011) Mechanotransduction: the role of mechanical stress, myocyte shape, and cytoskeletal architecture on cardiac function. Eur J Physiol 462: 89–104. |
| [34] |
Frank D, Frey N (2011) Cardiac Z-disc Signaling Network. J Biol Chem 286: 9897–9904. doi: 10.1074/jbc.R110.174268
|
| [35] |
Paluch EK, Nelson CM, Biais N, et al. (2015) Mechanotransduction: use the force(s). BMC Biology 13: 47. doi: 10.1186/s12915-015-0150-4
|
| [36] |
Ross RS (2004) Molecular and mechanical synergy: cross-talk between integrins and growth factor receptors. Cardiovascular Res 63: 381–390. doi: 10.1016/j.cardiores.2004.04.027
|
| [37] |
Vogel V, Sheetz MP (2009) Cell fate regulation by coupling mechanical cycles to biochemical signaling pathways. Curr Biol Cell Biol 21: 38–46. doi: 10.1016/j.ceb.2009.01.002
|
| [38] | Dupont S, Morsut L, Aragona M, et al. (2011) Role of YAP/TAZ in mechanotransduction. Nature 474: 179–185. |
| [39] |
Goldmann WH, Auernheimer V, Thievessen I, et al. (2013) Vinculin, cell mechanics and tumour cell invasion. Cell Biol Int 37: 397–405. doi: 10.1002/cbin.10064
|
| [40] |
Kaminski A, Fedorchak GR, Lammerding J (2014) The cellular mastermind(?)– Mechanotransduction and the nucleus. Progress in Molecular Biology and Translational Science 126: 157–203. doi: 10.1016/B978-0-12-394624-9.00007-5
|
| [41] |
Wang N, Tytell JD, Ingber DE (2009) Mechanotransduction at a distance: mechanically coupling the extracellular matrix with the nucleus. Nat Rev Mol Cell Biology 10: 75–82. doi: 10.1038/nrm2594
|
| [42] | Alenghat FJ, Ingber DE (2002) Mechanotransduction: All signals point to cytoskeleton, Matrix, and Integrins. Sci StKE 119: pe6. |
| [43] | Auernheimer V, Lautscham LA, Leidenberger M, et al. (2015) Vinculin phosphorylation at residues V100 and Y1065 is required for cellular force transmission. J Cell Sci 128: 3435–3443. |
| [44] |
Goldmann WH (2002) Mechanical aspects of cell shape regulation and signaling. Cell Biol Int 26: 313–317. doi: 10.1006/cbir.2002.0857
|
| [45] |
Janoštiak R, Brábek J, Auernheimer V, et al. (2014) CAS directly interacts with vinculin to control mechanosensing and focal adhesion dynamics. Cell Mol Life Sci 71: 727–44. doi: 10.1007/s00018-013-1450-x
|
| [46] |
Samarel AM (2005) Costameres, focal adhesions, and cardiomyocyte mechanotransduction. Am J Physiol Heart Circ Physiol 289: H2291–H2301. doi: 10.1152/ajpheart.00749.2005
|
| [47] |
Butcher DT, Alliston T, Weaver VM (2009) A tense situation: forcing tumour progression. Nature Rev. Cancer 9: 108–122. doi: 10.1038/nrc2544
|
| [48] | Goldmann WH (2012b) Mechanotransduction and focal adhesions. Cell Biol Int 36: 649–652. |
| [49] |
Vogel V, Sheetz MP (2006) Local force and geometry sensing regulate cell functions. Nat Rev Molecular Cell Biology 7: 265–275. doi: 10.1038/nrm1890
|
| [50] |
Wang HB, Dembo M, Hanks SK, et al. (2001) Focal adhesion kinase is involved in mechanosensing during fibroblast migration. Proc Nat Acad Sci USA 98: 11295–11300. doi: 10.1073/pnas.201201198
|
| [51] | Bendig G, Grimmler M, Huttner IG, et al. (2006) Integrin-linked kinase, a novel component of the cardiac mechanical stretch sensor, controls contractility in the zebrafish heart. Genes Dev 20: 2361–2372. |
| [52] |
Shih YRV, Tseng KF, Lai HY, et al. (2011) Matrix stiffness regulation of integrin-mediated mechanotransduction during osteogenic differentiation of human mesenchymal stem cells. J Bone Miner Res 26: 730–738. doi: 10.1002/jbmr.278
|
| [53] |
Mehta PK, Griendling KK (2006) Angiotensin II cell signaling: physiological and pathological effects in the cardiovascular system. Am J Physiol Cell Physiol 292: C82–C97. doi: 10.1152/ajpcell.00287.2006
|
| [54] | Schwartz MA, Assoian RK (2001) Integrins and cell proliferation: regulation of cyclin-dependent kinases via cytoplasmic signaling pathways. J Cell Sci 114: 2553–2560. |
| [55] | McBeath R, Pirone DM, Nelson CM, et al. (2004) Cell shape, cytoskeletal tension, and RhoA regulate stem cell lineage commitment. Cell 6: 483–495. |
| [56] |
Bertrand AT, Ziaei S, Ehret C, et al. (2014) Cellular microenvironments reveal defective mechanosensing responses and elevated YAP signaling in LMNA-mutated muscle precursors. J Cell Sci 127: 2873–2884. doi: 10.1242/jcs.144907
|
| [57] |
Yuan JM, Chyan AL, Zhou HX, et al. (2008) The effect of macromolecular crowding on the mechanical stability of protein molecules. Protein Sci 17: 2156–2166. doi: 10.1110/ps.037325.108
|
| [58] |
Ladoux B, Nelson WJ, Yan J, et al. (2015) The mechanotransduction machinery at work at adherens junctions. Integr Biol 7: 1109–1119. doi: 10.1039/C5IB00070J
|
| [59] |
Agrawal S, Agrawal A, Doughty B, et al. (2003) Cutting edge: different toll-like receptors agonists instruct dendritic cells to induce distinct responses via differential modulation of extracellular signal-regulated kinase-activated protein kinase and cFos. J Immunol 171: 4984–4989. doi: 10.4049/jimmunol.171.10.4984
|
| [60] | Chiu JJ, Chien S (2011) Effects of disturbed flow on vascular endothelium: pathophysiological basis and clinical perspectives. Physiol Rev 91: 10.1152. |
| [61] |
Maroto R, Raso A, Wood TG, et al. (2005) TRPC1 forms the stretch-activated cation channel in vertebrate cells. Nat Cell Biol 7: 179–185. doi: 10.1038/ncb1218
|
| [62] |
Davies PF, Tripathi SC (1993) Mechanical stress mechanisms and the cell: an endothelial paradigm. Cir Res 72: 239–245. doi: 10.1161/01.RES.72.2.239
|
| [63] | Burkholder TJ (2008) Mechanotransduction in skeletal muscle. Front Biosci 12: 174–191. |
| [64] |
Benavides DT, Egli M (2014) Calcium's Role in Mechanotransduction during muscle development. Cell Physiol Biochem 33: 249–272. doi: 10.1159/000356667
|
| [65] |
Schwartz MA, Simone DW (2008) Cell adhesion receptors in mechano-transduction. Curr Opion Cell Biol 20: 551–556. doi: 10.1016/j.ceb.2008.05.005
|
| [66] | Ingber DE (2003) Mechanobiology and diseases of mechanotransduction. Ann Med 35: 1–14. |
| [67] | Schreiner SM, Koo PK, Zhao Y, et al (2015) The tethering of chromatin to the nuclear envelope supports nuclear mechanics. Nat Comm 6: 7159. |
| [68] | Engler AJ, Kumar S (2014) Mechanosensation. Progress in Molecular Biology and Translational Science, Academic Press. 126: 1–384. |
| [69] |
Isermann P, Lammerding J (2013) Nuclear mechanics and mechanotransduction in health and disease. Curr Biol 23: R1113–1121. doi: 10.1016/j.cub.2013.11.009
|
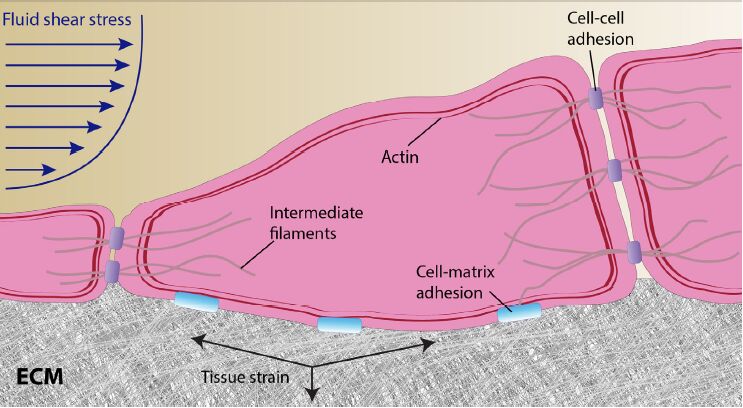
Figures(1)

José Luis Alonso, Wolfgang H. Goldmann. Cellular mechanotransduction[J]. AIMS Biophysics, 2016, 3(1): 50-62. doi: 10.3934/biophy.2016.1.50







 DownLoad:
DownLoad: