Citation: Roman Czapla, Vladimir V. Mityushev. A criterion of collective behavior of bacteria[J]. Mathematical Biosciences and Engineering, 2017, 14(1): 277-287. doi: 10.3934/mbe.2017018

| [1] | [ N. I. Akhiezer, Elements of Theory of Elliptic Functions Nauka, 1970 (in Russian); Engl. transl. AMS, 1990. |
| [2] | [ R. Czapla,V. V. Mityushev,W. Nawalaniec, Effective conductivity of random two-dimensional composites with circular non-overlapping inclusions, Computational Materials Science, 63 (2012): 118-126. |
| [3] | [ R. Czapla,V. V. Mityushev,W. Nawalaniec, Simulation of representative volume elements for random 2D composites with circular non-overlapping inclusions, Theoretical and Applied Informatics, 24 (2012): 227-242. |
| [4] | [ R. Czapla,V. V. Mityushev,N. Rylko, Conformal mapping of circular multiply connected domains onto segment domains, Electron. Trans. Numer. Anal., 39 (2012): 286-297. |
| [5] | [ S. Gluzman,D. A. Karpeev,L. V. Berlyand, Effective viscosity of puller-like microswimmers: A renormalization approach, J. R. Soc. Interface, 10 (2013): 1-10. |
| [6] | [ V. V. Mityushev, Representative cell in mechanics of composites and generalized Eisenstein--Rayleigh sums, Complex Variables, 51 (2006): 1033-1045. |
| [7] | [ V. V. Mityushev,P. Adler, Longitudial permeability of a doubly periodic rectangular array of circular cylinders, I, ZAMM (Journal of Applied Mathematics and Mechanics), 82 (2002): 335-345. |
| [8] | [ V. V. Mityushev,W. Nawalaniec, Basic sums and their random dynamic changes in description of microstructure of 2D composites, Computational Materials Science, 97 (2015): 64-74. |
| [9] | [ V. V. Mityushev,N. Rylko, Optimal distribution of the non-overlapping conducting disks, Multiscale Model. Simul., 10 (2012): 180-190. |
| [10] | [ W. Nawalaniec, Algorithms for computing symbolic representations of basic e–sums and their application to composites Journal of Symbolic Computation 74 (2016), 328–345. |
| [11] | [ M. Potomkin, V. Gyrya, I. Aranson and L. Berlyand, Collision of microswimmers in viscous fluid Physical Review E 87 (2013), 053005. |
| [12] | [ S. D. Ryan,L. Berlyand,B. M. Haines,D. A. Karpeev, A kinetic model for semi-dilute bacterial suspensions, Multiscale Model. Simul., 11 (2013): 1176-1196. |
| [13] | [ S. D. Rayn, B. M. Haines, L. Berlyand, F. Ziebert and I. S. Aranson, Viscosity of bacterial suspensions: Hydrodynamic interactions and self-induced noise Rapid Communication to Phys. Rev. E 83 (2011), 050904(R). |
| [14] | [ S. D. Ryan, A. Sokolov, L. Berlyand and I. S. Aranson, Correlation properties of collective motion in bacterial suspensions New Journal of Physics 15 (2013), 105021, 18pp. |
| [15] | [ N. Rylko, Representative volume element in 2D for disks and in 3D for balls, J. Mechanics of Materials and Structures, 9 (2014): 427-439. |
| [16] | [ A. Sokolov and I. S. Aranson, Reduction of viscosity in suspension of swimming bacteria Phys. Rev. Lett. 103 (2009), 148101. |
| [17] | [ A. Sokolov and I. S. Aranson, Physical properties of collective motion in suspensions of bacteria Phys. Rev. Lett. 109 (2012), 248109. |
| [18] | [ A. Sokolov, I. S. Aranson, J. O. Kessler and R. E. Goldstein, Concentration dependence of the collective dynamics of swimming bacteria Physical Review Letters 98 (2007), 158102. |
| [19] | [ A. Sokolov, R. E. Goldstein, F. I. Feldstein and I. S. Aranson, Enhanced mixing and spatial instability in concentrated bacteria suspensions, Phys. Rev. E, 80 (2009), 031903. |
| [20] | [ M. Tournus,L. V. Berlyand,A. Kirshtein,I. Aranson, Flexibility of bacterial flagella in external shear results in complex swimming trajectories, Journal of the Royal Society Interface, 12 (2015): 1-11. |
| [21] | [ A. Weil, Elliptic Functions According to Eisenstein and Kronecker Springer-Verlag, 1976. |
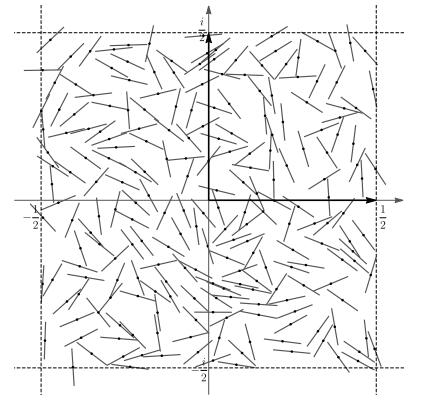
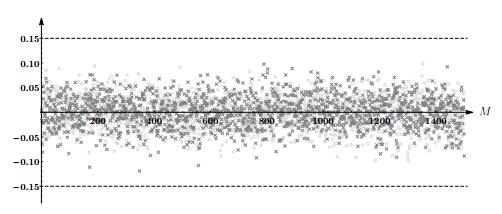
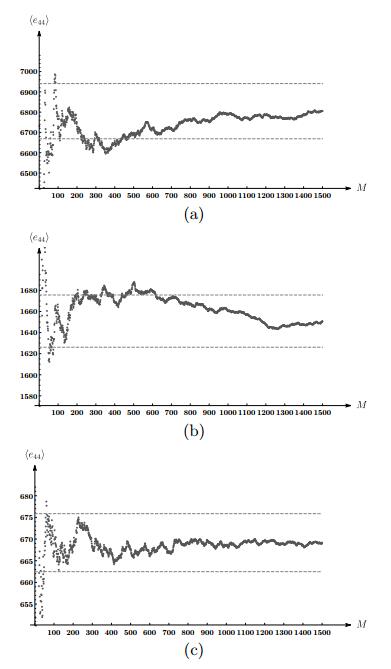

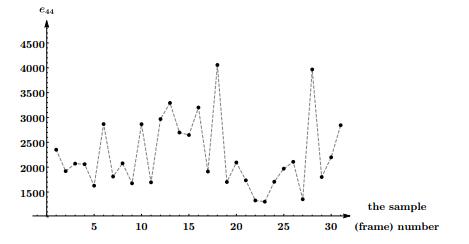
Figures(5) / Tables(4)

Roman Czapla, Vladimir V. Mityushev. A criterion of collective behavior of bacteria[J]. Mathematical Biosciences and Engineering, 2017, 14(1): 277-287. doi: 10.3934/mbe.2017018







 DownLoad:
DownLoad: