Citation: Miguel de Simón-Martín, Montserrat Díez-Mediavilla, Cristina Alonso-Tristán. Real Energy Payback Time and Carbon Footprint of a GCPVS[J]. AIMS Energy, 2017, 5(1): 77-95. doi: 10.3934/energy.2017.1.77

| [1] | Perpiñán O, Lorenzo E, Castro M A, et al. (2009) Energy payback time of grid connected PV systems: comparison between tracking and fixed systems. Prog Photovoltaics 17(2): 137–147. |
| [2] | Fthenakis V, Alsema E A, de Wild-Scholten M (2005) Life cycle assessment of photovoltaics: perceptions, needs, and challenges, in: Conference Record of the Thirty-first IEEE Photovoltaic Specialists Conference, 1655–1658. |
| [3] | Keoleian G A, Lewis G M (1997) Application of life-cycle energy analysis to photovoltaic module design. Prog Photovoltaics 5(4): 287–300. |
| [4] | Sherwani A, Usmani J, Varun (2010) Life cycle assessment of solar PV based electricity generation systems: A review. Renew Sust Energ Rev 14(1): 540–544. |
| [5] | Tahara K, Kojima T, Inaba A (1997) Estimation of power plants by LCA. Kagaku Kogaku Ronbun 23(1): 93–94. |
| [6] | de Wild-Scholten M, Alsema E (2004) Towards cleaner solar PV: Environmental and health impacts of crystalline silicon photovoltaics. Refocus 5(5): 46–49. |
| [7] | Fthenakis V, Alsema E (2006) Photovoltaics energy payback times, greenhouse gas emissions and external costs: 2004 – early 2005 status. Prog Photovoltaics 14(3): 275–280. |
| [8] | Bayod-Rújula A A, Lorente-Lafuente A M, Cirez-Oto F (2011) Environmental assessment of grid connected photovoltaic plants with 2-axis tracking versus fixed modules systems. Energy 36(5): 3148–3158. |
| [9] | Mason J E, Fthenakis V M, Hansen T, et al. (2006) Energy payback and life-cycle CO2 emissions of the BOS in an optimized 3.5 MW PV installation. Prog Photovoltaics 14(2): 179–190. |
| [10] | Nawaz I, Tiwari G (2006) Embodied energy analysis of photovoltaic (PV) system based on macroand micro-level. Energ Policy 34(17): 3144–3152. |
| [11] | SoDa Team SoDa: HelioClim-3, 2016, Available from: http://www.soda-pro.com/web-services/radiation/helioclim-3-for-free. |
| [12] | Europan Commission: PVGIS- PV Potential Estimation Utility, 2016, Available from: http://re.jrc.ec.europa.eu/pvgis/apps4/pvest.php. |
| [13] | Hacke P, Smith R, Terwilliger K, et al. (2013) Testing and Analysis for Lifetime Prediction of Crystalline Silicon PV Modules Undergoing Degradation by System Voltage Stress. IEEE J Photovoltaics 3(1): 246–253. |
| [14] | Muñoz M A, Alonso-García M C, Vela N, et al. (2011) Early degradation of silicon PV modules and guaranty conditions. Sol Energy 85(9): 2264–2274. |
| [15] | Jordan D C, Kurtz S R (2012) Photovoltaic Degradation Rates-An Analytical Review. NREL/JA- 5200-51664 1(1): 1–32. |
| [16] | Osterwald C R, Anderberg A, Rummel S, et al. (2002) Degradation analysis of weathered crystalline-silicon PV modules, in: Conference Record of the Twenty-Ninth IEEE Photovoltaic Specialists Conference, 1392–1395. |
| [17] | University of Manchester: Carbon calculations over the life cycle of industrial activities, 2016, Available from:http://www.ccalc.org.uk/. |
| [18] | Postnote from the Parliamentary Office of Science and Technology: Carbon footprint of electricity generation, 2006, 268:1–4. |
| [19] | Díez-Mediavilla M, Alonso-Tristán C, Rodríguez-Amigo M, et al. (2012) Performance analysis of PV plants: Optimization for improving profitability. Energ Convers Manage 54(1): 17–23. |
| [20] | Khasawneh Q A, Damra Q A, Salman O H B Determining the Optimum Tilt Angle for Solar Applications in Northern Jordan 9(3): 187–193. |
| [21] | Skeiker K Optimum tilt angle and orientation for solar collectors in Syria 50(1): 2439–2448. |
| [22] | Government of Spain: Carbon emission factors and primary energy conversion coe cients for the different electrical energy sources in the Building Sector in Spain, 2014, IDAE, 1–32. |
| [23] | Alsema E (1998) Energy Requirements and CO2 Mitigation Potential of PV Systems, in: BNL/NREL Workshop PV and the Environment, 1–11. |
| [24] | Adelstein J, Sekulic B (2005) Performance and reliability of a 1-kW amorphous silicon photovoltaic roofing system, in: Conference Record of the Thirty-first IEEE Photovoltaic Specialists Conference, 1627–1630. |
| [25] | Chamberlin C E, Rocheleau M A, Marshall M W, et al. (2011) Comparison of PV module performance before and after 11 and 20 years of field exposure, in: 2011 37th IEEE Photovoltaic Specialists Conference (PVSC), 101–105. |
| [26] | Lorenzo E, Zilles R, Moretón R, et al. (2013) Performance analysis of a 7-kW crystalline silicon generator after 17 years of operation in Madrid. Prog Photovoltaics 22(12): 1273–1279. |
| [27] | Espinosa N, García-Valverde R, Urbina A, et al. (2011) A life cycle analysis of polymer solar cell modules prepared using roll-to-roll methods under ambient conditions. Sol Energ Mat Sol C 95(5): 1293–1302. |
| [28] | Fthenakis V M, Kim H C (2013) Life cycle assessment of high-concentration photovoltaic systems. Prog Photovoltaics 21(3): 379–388. |
| [29] | Hondo H (2005) Life cycle GHG emission analysis of power generation systems: Japanese case. Energy 30(11-12): 2042–2056. |
| [30] |
Peng J, Lu L, Yang H (2013) Review on life cycle assessment of energy payback and greenhouse gas emission of solar photovoltaic systems. Renew Sust Energ Rev 19: 255–274. doi: 10.1016/j.rser.2012.11.035

|
| [31] | Jiao Y, Salce A, Ben W, et al. (2011) Siemens and siemens-like processes for producing photovoltaics: Energy payback time and lifetime carbon emissions. JOM 63(1): 28–31. |
| [32] | Mann S A, de Wild-Scholten M J, Fthenakis V M, et al. (2013) The energy payback time of advanced crystalline silicon PV modules in 2020: A prospective study. Prog Photovoltaics 22(11): 1180–1194. |
| [33] | Mohr N J, Meijer A, Huijbregts M a J, et al. (2013) Environmental life cycle assessment of roof-integrated flexible amorphous silicon/nanocrystalline silicon solar cell laminate. Prog Photovoltaics 21(4): 802–815. |
| [34] | Yamada K, Komiyama H, Kato K, et al. (1995) Evaluation of photovoltaic energy systems in terms of economics, energy and CO2 emissions. Energ Convers Manage 36(6–9): 819–822. |
| [35] | Hammond G, Jones C (2008) Embodied energy and carbon in construction materials. P I Civil Eng Energ 161(2): 87–98. |
| [36] | Pacca S, Sivaraman D, Keoleian G A (2007) Parameters affecting the life cycle performance of PV technologies and systems. Energ Policy 35(6): 3316–3326. |
| [37] | European Comission: Eurostat database, 2016, Available from: http://ec.europa.eu/ eurostat/data/database. |
| [38] | Davis S C, Diegel S W, Boundy R G (2012) Transportation Energy Data Book, volume 1, 31st edition, Oak Laboratory: U.S. Department of Energy. |
| [39] | European Environment Agency E U (2016) Explaining road transport emissions, volume 1, 1st edition, Luxembourg: Publications O ce of the European Union. |
| [40] | Oficina Catalana del Canvi Climátic E S (2011) Practical Guide for the Carbon Emissions Calculation. Oficina Catalana del Canvi Climátic 1(1): 1–32. |
| [41] | Joint Research Centre I E T (Ed.) (2013) Well-to-wheels analysis of future automotive fuels and powertrains in the European context, volume 1, 1st edition, Luxembourg: Publications Office of the European Union. |
| [42] | Pucker N, Schappacher W (1994) Installation of new energy systems: Energy balances and installation times; application to a photovoltaic system. Renew Energ 5(1–4): 212–214. |
| [43] | Previ A, Iliceto A, Belli G, et al. The 3.3 MW-peak photovoltaic power station at Serre, in: Proceedings of 1994 IEEE 1st World Conference on Photovoltaic Energy Conversion - WCPEC (A Joint Conference of PVSC, PVSEC and PSEC), volume 1, 750–753. |
| [44] | Iliceto A, Vigotti R (1998) The largest PV installation in Europe: Perspectives of multimegawatt PV. Renew Energ 15(1): 48–53. |

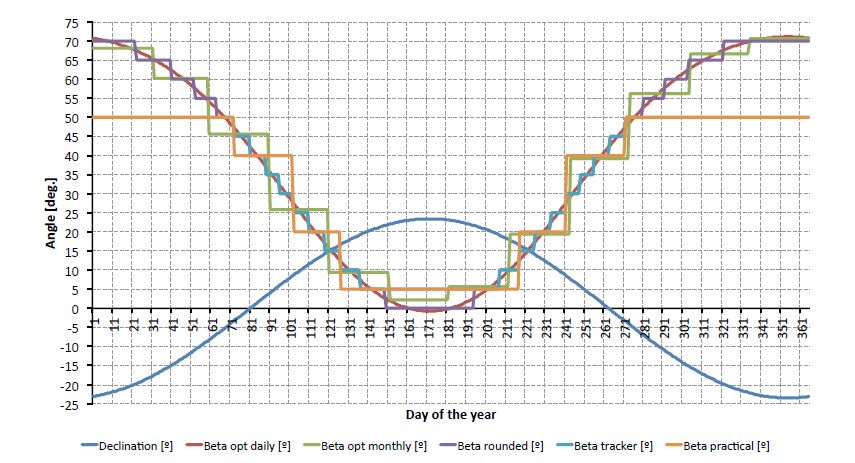

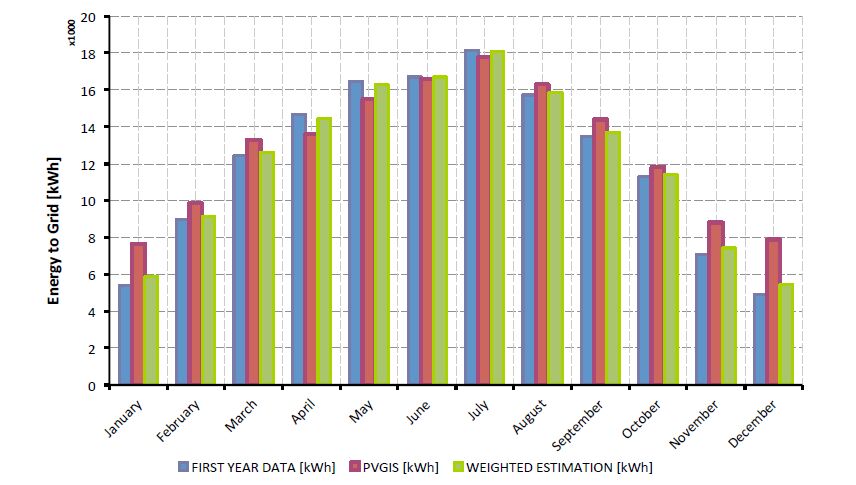
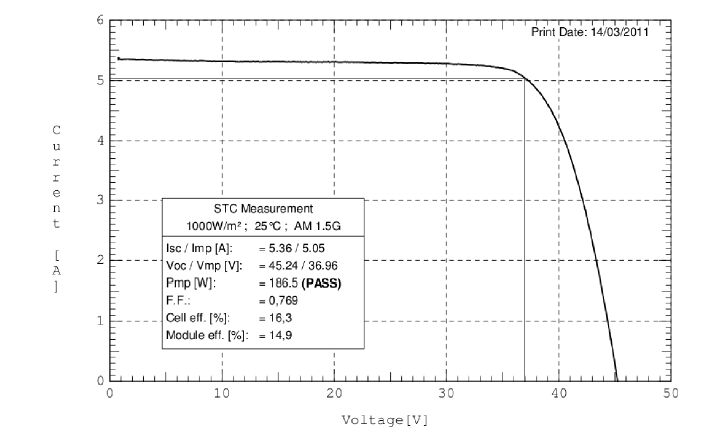
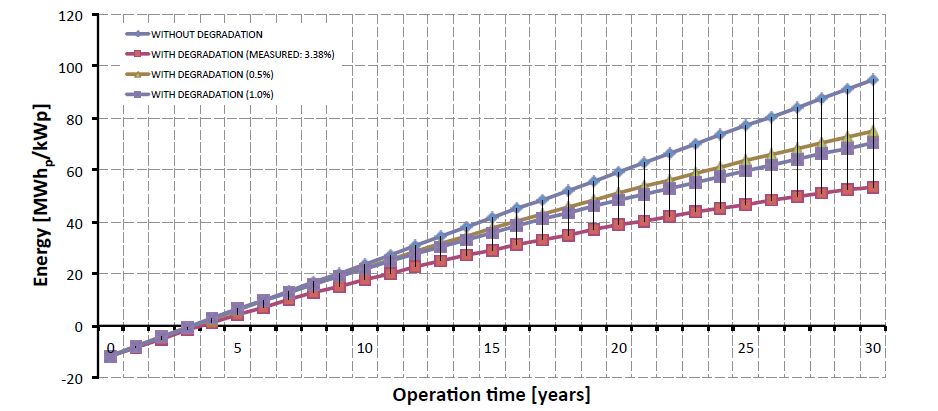

Figures(7) / Tables(6)

Miguel de Simón-Martín, Montserrat Díez-Mediavilla, Cristina Alonso-Tristán. Real Energy Payback Time and Carbon Footprint of a GCPVS[J]. AIMS Energy, 2017, 5(1): 77-95. doi: 10.3934/energy.2017.1.77







 DownLoad:
DownLoad: