We study the mathematical properties of a model of cell division structured by two variables – the size and the size increment – in the case of a linear growth rate and a self-similar fragmentation kernel. We first show that one can construct a solution to the related two dimensional eigenproblem associated to the eigenvalue $ 1 $ from a solution of a certain one dimensional fixed point problem. Then we prove the existence and uniqueness of this fixed point in the appropriate $ {\rm{L}} ^1 $ weighted space under general hypotheses on the division rate. Knowing such an eigenfunction proves useful as a first step in studying the long time asymptotic behaviour of the Cauchy problem.
Citation: Pierre Gabriel, Hugo Martin. Steady distribution of the incremental model for bacteria proliferation[J]. Networks and Heterogeneous Media, 2019, 14(1): 149-171. doi: 10.3934/nhm.2019008
We study the mathematical properties of a model of cell division structured by two variables – the size and the size increment – in the case of a linear growth rate and a self-similar fragmentation kernel. We first show that one can construct a solution to the related two dimensional eigenproblem associated to the eigenvalue $ 1 $ from a solution of a certain one dimensional fixed point problem. Then we prove the existence and uniqueness of this fixed point in the appropriate $ {\rm{L}} ^1 $ weighted space under general hypotheses on the division rate. Knowing such an eigenfunction proves useful as a first step in studying the long time asymptotic behaviour of the Cauchy problem.

| [1] | Cell growth and division: Ⅰ. a mathematical model with applications to cell volume distributions in mammalian suspension cultures. Biophysical Journal (1967) 7: 329-351. |
| [2] |
E. Bernard, M. Doumic and P. Gabriel, Cyclic asymptotic behaviour of a population reproducing by fission into two equal parts, Kinetic Related Models, to appear, arXiv: 1609.03846. |
| [3] |
H. Brezis, Functional Analysis, Sobolev Spaces and Partial Differential Equations, Universitext. Springer, New York, 2011. |
| [4] |
M. J. Cáceres, J. A. Cañizo and S. Mischler, Rate of convergence to an asymptotic profile for the self-similar fragmentation and growth-fragmentation equations, J. Math. Pures Appl. (9), 96 (2011), 334–362. |
| [5] |
Irreducible compact operators. Math. Z. (1986) 192: 149-153. 
|
| [6] |
Analysis of a population model structured by the cells molecular content. Math. Model. Nat. Phenom. (2007) 2: 121-152.
|
| [7] |
Statistical estimation of a growth-fragmentation model observed on a genealogical tree. Bernoulli (2015) 21: 1760-1799.
|
| [8] |
Eigenelements of a general aggregation-fragmentation model. Math. Models Methods Appl. Sci. (2010) 20: 757-783.
|
| [9] |
Y. Du, Order structure and topological methods in nonlinear partial differential equations, Vol. 1. Maximum principles and applications, Series in Partial Differential Equations and Applications. World Scientific Publishing Co. Pte. Ltd., Hackensack, NJ, 2006. Maximum principles and applications. |
| [10] |
Steady size distributions for cells in one-dimensional plant tissues. J. Math. Biol. (1991) 30: 101-123.
|
| [11] |
Sur un point de la théorie des fonctions géné ratrices d'Abel. Acta Math. (1903) 27: 339-351.
|
| [12] |
J. A. J. Metz and O. Diekmann, editors, The Dynamics of Physiologically Structured Populations, volume 68 of Lecture Notes in Biomathematics, Springer-Verlag, Berlin, 1986. |
| [13] |
General entropy equations for structured population models and scattering. C. R., Math., Acad. Sci. Paris (2004) 338: 697-702.
|
| [14] |
P. Michel, S. Mischler and B. Perthame, General relative entropy inequality: An illustration on growth models, J. Math. Pures Appl. (9), 84 (2005), 1235–1260. |
| [15] |
Stability in a nonlinear population maturation model. Math. Models Methods Appl. Sci. (2002) 12: 1751-1772.
|
| [16] |
Spectral analysis of semigroups and growth-fragmentation equations. Ann. Inst. Henri Poincaré, Anal. Non Linéaire (2016) 33: 849-898.
|
| [17] |
How does variability in cells aging and growth rates influence the malthus parameter?. Kinetic and Related Models (2017) 10: 481-512.
|
| [18] |
B. Perthame, Transport Equations in Biology, Frontiers in Mathematics. Birkhäuser Verlag, Basel, 2007. |
| [19] |
Exponential decay for the fragmentation or cell-division equation. J. Differential Equations (2005) 210: 155-177.
|
| [20] |
J. D. F. Richard L. Burden, Numerical Analysis, Boston, MA: PWS Publishing Company; London: ITP International Thomson Publishing, 5th ed. edition, 1993. |
| [21] |
Transport theory for growing cell populations. J. Theoret. Biol. (1983) 103: 181-199.
|
| [22] |
Adder and a coarse-grained approach to cell size homeostasis in bacteria. Current Opinion in Cell Biology (2016) 38: 38-44.
|
| [23] |
H. H. Schaefer, Banach Lattices and Positive Operators, Springer-Verlag, New York-Heidelberg, 1974. |
| [24] |
A new model for age-size structure of a population. Ecology (1967) 48: 910-918.
|
| [25] | Cell-size control and homeostasis in bacteria. Current Biology (2015) 25: 385-391. |
| [26] |
Dynamics of populations structured by internal variables. Math. Z. (1985) 189: 319-335.
|
| [27] |
G. F. Webb, Theory of Nonlinear Age-Dependent Population Dynamics, volume 89 of Monographs and Textbooks in Pure and Applied Mathematics, Marcel Dekker, Inc., New York, 1985. |
| [28] |
An operator-theoretic formulation of asynchronous exponential growth. Trans. Amer. Math. Soc. (1987) 303: 751-763.
|
Figures(3)
Pierre Gabriel, Hugo Martin. Steady distribution of the incremental model for bacteria proliferation[J]. Networks and Heterogeneous Media, 2019, 14(1): 149-171. doi: 10.3934/nhm.2019008

schematic representation of the variables on an E. coli bacterium
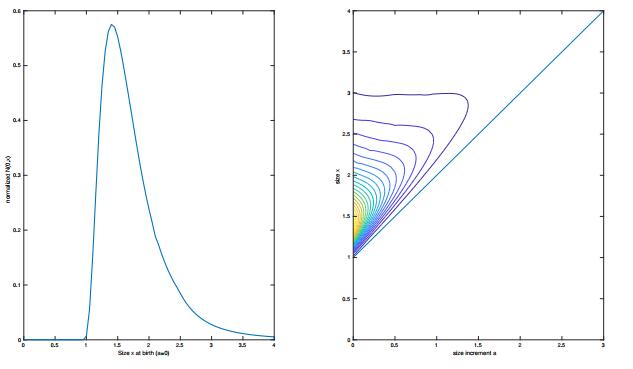
Left: simulation of the function
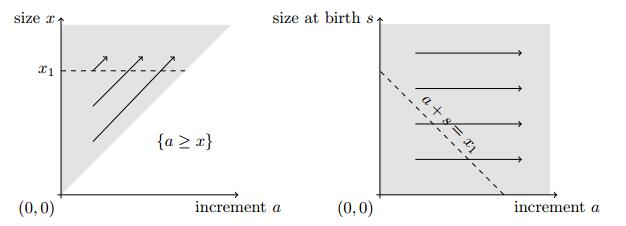
Domain of the model, with respect to the choice of variables to describe the bacterium. Grey: domain where the bacteria densities may be positive. Arrows: transport. Left: size increment/size. Right: size increment/birth size. Dashed: location of cells of size $x_1$






 DownLoad:
DownLoad: