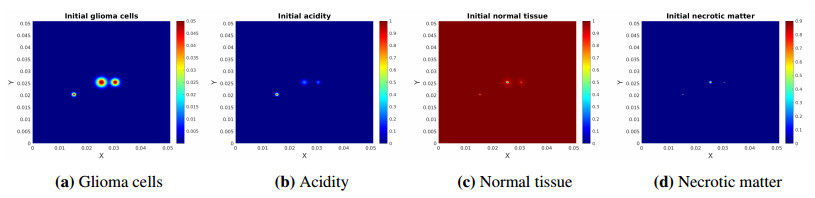
We propose a multiphase modeling approach to describe glioma pseudopalisade patterning under the influence of acidosis. The phases considered at the model onset are glioma, normal tissue, necrotic matter, and interstitial fluid in a void-free volume with acidity represented by proton concentration. We start from mass and momentum balance to characterize the respective volume fractions and deduce reaction-cross diffusion equations for the space-time evolution of glioma, normal tissue, and necrosis. These are supplemented with a reaction-diffusion equation for the acidity dynamics and lead to formation of patterns which are typical for high grade gliomas. Unlike previous works, our deduction also works in higher dimensions and involves less restrictions. We also investigate the existence of weak solutions to the obtained system of equations and perform numerical simulations to illustrate the solution behavior and the pattern occurrence.
Citation: Pawan Kumar, Christina Surulescu, Anna Zhigun. Multiphase modelling of glioma pseudopalisading under acidosis[J]. Mathematics in Engineering, 2022, 4(6): 1-28. doi: 10.3934/mine.2022049
We propose a multiphase modeling approach to describe glioma pseudopalisade patterning under the influence of acidosis. The phases considered at the model onset are glioma, normal tissue, necrotic matter, and interstitial fluid in a void-free volume with acidity represented by proton concentration. We start from mass and momentum balance to characterize the respective volume fractions and deduce reaction-cross diffusion equations for the space-time evolution of glioma, normal tissue, and necrosis. These are supplemented with a reaction-diffusion equation for the acidity dynamics and lead to formation of patterns which are typical for high grade gliomas. Unlike previous works, our deduction also works in higher dimensions and involves less restrictions. We also investigate the existence of weak solutions to the obtained system of equations and perform numerical simulations to illustrate the solution behavior and the pattern occurrence.

| [1] |
J. Alfonso, A. Köhn-Luque, T. Stylianopoulos, F. Feuerhake, A. Deutsch, H. Hatzikirou, Why one-size-fits-all vaso-modulatory interventions fail to control glioma invasion: in silico insights, Sci. Rep., 6 (2016), 37283. doi: 10.1038/srep37283

|
| [2] |
R. J. Atkin, R. E. Craine, {Continuum theories of mixtures: basic theory and historical development}, The Quarterly Journal of Mechanics and Applied Mathematics, 29 (1976), 209–244. doi: 10.1093/qjmam/29.2.209
|
| [3] |
C. Barbarosie, Representation of divergence-free vector fields, Q. Appl. Math., 69 (2011), 309–316. doi: 10.1090/S0033-569X-2011-01215-2
|
| [4] | N. Bellomo, Modeling complex living systems, Boston: Birkhäuser, 2008. |
| [5] |
D. J. Brat, A. A. Castellano-Sanchez, S. B. Hunter, M. Pecot, C. Cohen, E. H. Hammond, et al., Pseudopalisades in glioblastoma are hypoxic, express extracellular matrix proteases, and are formed by an actively migrating cell population, Cancer Res., 64 (2004), 920–927. doi: 10.1158/0008-5472.CAN-03-2073
|
| [6] |
D. J. Brat, T. B. Mapstone, Malignant glioma physiology: Cellular response to hypoxia and its role in tumor progression, Ann. Intern. Med., 138 (2003), 659–668. doi: 10.7326/0003-4819-138-8-200304150-00014
|
| [7] |
C. Breward, H. Byrne, C. Lewis, The role of cell-cell interactions in a two-phase model for avascular tumour growth, J. Math. Biol., 45 (2002), 125–152. doi: 10.1007/s002850200149
|
| [8] |
H. Byrne, J. King, D. McElwain, L. Preziosi, A two-phase model of solid tumour growth, Appl. Math. Lett., 16 (2003), 567–573. doi: 10.1016/S0893-9659(03)00038-7
|
| [9] | M. Conte, C. Surulescu, Mathematical modeling of glioma invasion: acid- and vasculature mediated go-or-grow dichotomy and the influence of tissue anisotropy, Appl. Math. Comput., 407 (2021), 126305. |
| [10] |
G. Corbin, A. Klar, C. Surulescu, C. Engwer, M. Wenske, J. Nieto, et al., Modeling glioma invasion with anisotropy- and hypoxia-triggered motility enhancement: From subcellular dynamics to macroscopic pdes with multiple taxis, Math. Models Methods Appl. Sci., 31, (2021), 177–222. doi: 10.1142/S0218202521500056
|
| [11] | A. Dietrich, N. Kolbe, N. Sfakianakis, C. Surulescu, Multiscale modeling of glioma invasion: from receptor binding to flux-limited macroscopic pdes, 2020, arXiv: 2010.03277. |
| [12] | D. A. Drew, S. L. Passman, Theory of multicomponent fluids, New York: Springer, 1999. |
| [13] |
S. E. Eikenberry, T. Sankar, M. C. Preul, E. J. Kostelich, C. J. Thalhauser, Y. Kuang, Virtual glioblastoma: growth, migration and treatment in a three-dimensional mathematical model, Cell Prolif., 42 (2009), 511–528. doi: 10.1111/j.1365-2184.2009.00613.x
|
| [14] |
C. Engwer, T. Hillen, M. Knappitsch, C. Surulescu, Glioma follow white matter tracts: a multiscale DTI-based model, J. Math. Biol., 71 (2015), 551–582. doi: 10.1007/s00285-014-0822-7
|
| [15] | C. Engwer, A. Hunt, C. Surulescu, Effective equations for anisotropic glioma spread with proliferation: a multiscale approach and comparisons with previous settings, Math. Med. Biol., 33 (2015), 435–459. |
| [16] |
C. Engwer, M. Knappitsch, C. Surulescu, A multiscale model for glioma spread including cell-tissue interactions and proliferation, Math. Biosci. Eng., 13 (2016), 443–460. doi: 10.3934/mbe.2015011
|
| [17] |
H. Garcke, K. F. Lam, R. Nürnberg, E. Sitka, A multiphase cahn-hilliard-darcy model for tumour growth with necrosis, Math. Models Methods Appl. Sci., 28 (2018), 525–577. doi: 10.1142/S0218202518500148
|
| [18] | A. Hunt, C. Surulescu, A multiscale modeling approach to glioma invasion with therapy, Vietnam J. Math., 45 (2016), 221–240. |
| [19] |
T. L. Jackson, H. M. Byrne, A mechanical model of tumor encapsulation and transcapsular spread, Math. Biosci., 180 (2002), 307–328. doi: 10.1016/S0025-5564(02)00118-9
|
| [20] |
A. F. Jones, H. M. Byrne, J. S. Gibson, J. W. Dold, A mathematical model of the stress induced during avascular tumour growth, J. Math. Biol., 40 (2000), 473–499. doi: 10.1007/s002850000033
|
| [21] |
A. Jüngel, The boundedness-by-entropy method for cross-diffusion systems, Nonlinearity, 28 (2015), 1963–2001. doi: 10.1088/0951-7715/28/6/1963
|
| [22] |
A. Jüngel, I. V. Stelzer, Entropy structure of a cross-diffusion tumor-growth model, Math. Models Methods Appl. Sci., 22 (2012), 1250009. doi: 10.1142/S0218202512500091
|
| [23] |
P. Kleihues, F. Soylemezoglu, B. Schäuble, B. W. Scheithauer, P. C. Burger, Histopathology, classification, and grading of gliomas, Glia, 15 (1995), 211–221. doi: 10.1002/glia.440150303
|
| [24] | N. Kolbe, N. Sfakianakis, C. Stinner, C. Surulescu, J. Lenz, Modeling multiple taxis: Tumor invasion with phenotypic heterogeneity, haptotaxis, and unilateral interspecies repellence, Discrete Contin. Dyn. Syst. Ser. B, 26 (2021), 443–481. |
| [25] | P. Kumar, Mathematical modeling of glioma patterns as a consequence of acidosis and hypoxia, PhD thesis of TU Kaiserslautern, 2021. |
| [26] |
P. Kumar, J. Li, C. Surulescu, Multiscale modeling of glioma pseudopalisades: contributions from the tumor microenvironment, J. Math. Biol., 82 (2021), 49. doi: 10.1007/s00285-021-01599-x
|
| [27] |
P. Kumar, C. Surulescu, A flux-limited model for glioma patterning with hypoxia-induced angiogenesis, Symmetry, 12 (2020), 1870. doi: 10.3390/sym12111870
|
| [28] |
G. Lemon, J. King, Multiphase modelling of cell behaviour on artificial scaffolds: effects of nutrient depletion and spatially nonuniform porosity, Math. Med. Biol., 24 (2007), 57–83. doi: 10.1093/imammb/dql020
|
| [29] | G. R. Martin, R. K. Jain, Noninvasive measurement of interstitial pH profiles in normal and neoplastic tissue using fluorescence ratio imaging microscopy, Cancer Res., 54 (1994), 5670–5674. |
| [30] |
A. Martínez-González, G. F. Calvo, L. A. Pérez Romasanta, V. M. Pérez-García, Hypoxic cell waves around necrotic cores in glioblastoma: a biomathematical model and its therapeutic implications, Bull. Math. Biol., 74 (2012), 2875–2896. doi: 10.1007/s11538-012-9786-1
|
| [31] |
K. J. Painter, T. Hillen, Mathematical modelling of glioma growth: the use of diffusion tensor imaging (DTI) data to predict the anisotropic pathways of cancer invasion, J. Theor. Biol., 323 (2013), 25–39. doi: 10.1016/j.jtbi.2013.01.014
|
| [32] |
L. Preziosi, A. Tosin, Multiphase modelling of tumour growth and extracellular matrix interaction: mathematical tools and applications, J. Math. Biol., 58 (2009), 625. doi: 10.1007/s00285-008-0218-7
|
| [33] |
L. Preziosi, G. Vitale, A multiphase model of tumor and tissue growth including cell adhesion and plastic reorganization, Math. Models Methods Appl. Sci., 21 (2011), 1901–1932. doi: 10.1142/S0218202511005593
|
| [34] |
G. Sciumè, S. Shelton, W. G. Gray, C. T. Miller, F. Hussain, M. Ferrari, et al., A multiphase model for three-dimensional tumor growth, New J. Phys., 15 (2013), 015005. doi: 10.1088/1367-2630/15/1/015005
|
| [35] |
A. M. Stein, T. Demuth, D. Mobley, M. Berens, L. M. Sander, A mathematical model of glioblastoma tumor spheroid invasion in a three-dimensional in vitro experiment, Biophys. J., 92 (2007), 356–365. doi: 10.1529/biophysj.106.093468
|
| [36] |
A. Tosin, L. Preziosi, Multiphase modeling of tumor growth with matrix remodeling and fibrosis, Math. Comput. Model., 52 (2010), 969–976. doi: 10.1016/j.mcm.2010.01.015
|
| [37] |
J. O. Waldeland, S. Evje, A multiphase model for exploring tumor cell migration driven by autologous chemotaxis, Chem. Eng. Sci., 191 (2018), 268–287. doi: 10.1016/j.ces.2018.06.076
|
| [38] |
B. A. Webb, M. Chimenti, M. P. Jacobson, D. L. Barber, Dysregulated pH: a perfect storm for cancer progression, Nat. Rev. Cancer, 11 (2011), 671–677. doi: 10.1038/nrc3110
|
| [39] | F. J. Wippold, M. Lämmle, F. Anatelli, J. Lennerz, A. Perry, Neuropathology for the neuroradiologist: Palisades and pseudopalisades, AJNR Am. J. Neuroradiol., 27 (2006), 2037–2041. |
| [40] |
S. M. Wise, J. S. Lowengrub, H. B. Frieboes, V. Cristini, Three-dimensional multispecies nonlinear tumor growth—Ⅰ: Model and numerical method, J. Theor. Biol., 253 (2008), 524–543. doi: 10.1016/j.jtbi.2008.03.027
|




Figures(5) / Tables(1)

Pawan Kumar, Christina Surulescu, Anna Zhigun. Multiphase modelling of glioma pseudopalisading under acidosis[J]. Mathematics in Engineering, 2022, 4(6): 1-28. doi: 10.3934/mine.2022049







 DownLoad:
DownLoad: