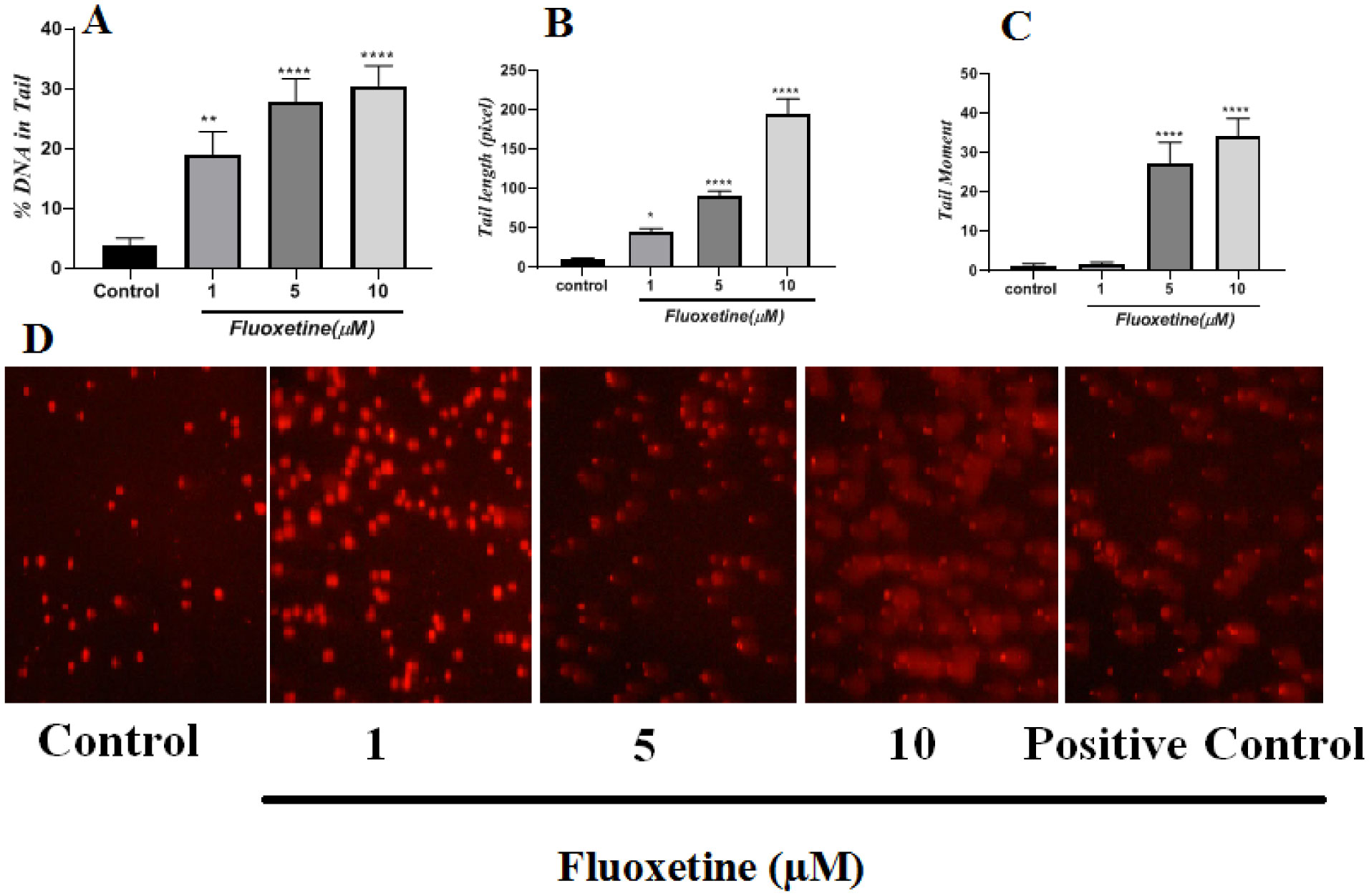
Fluoxetine is a selective serotonin reuptake inhibitor that is a commonly used drug for the treatment of depression and obsessive-compulsive disorders. Despite the positive effects of this drug, it seems to be associated with various side effects. Genotoxicity or DNA damage is an important side effect of some kinds of drugs. To date, the genotoxicity and cytotoxicity of fluoxetine are partially unknown. In the present study, some oxidative stress methods were used, such as ROS, MDA and GSH evaluation methods in HepG2 cells treated with fluoxetine (1–10 µM). A comet assay was used to evaluate the genotoxic effects of fluoxetine, and flow cytometry was used for apoptosis detection in these hepatic cells. Our data have shown that fluoxetine increased MDA and intracellular concentration of ROS significantly (P < 0.001), while the amount of GSH was reduced significantly (P < 0.001). Our results also indicated that fluoxetine increased the DNA damage of HepG2 cells. The tail percentage of DNA for control cells was 4%, but this percentage was 19%, 28% and 32% for 1, 5 and 10 µM of fluoxetine concentration, respectively (P < 0.01 and P < 0.001). The flow cytometry results have also shown increases in early and late apoptosis for fluoxetine (13.31% and 9.54%, respectively). In conclusion, the present study has shown that fluoxetine is able to induce oxidative stress-dependent DNA damage. Anyway, more studies are needed to accurately explore the molecular and cellular aspects of fluoxetine.
Citation: Somayeh Boshtam, Mohammad Shokrzadeh, Nasrin Ghassemi-Barghi. Fluoxetine induces oxidative stress-dependent DNA damage in human hepatoma cells[J]. AIMS Medical Science, 2023, 10(1): 69-79. doi: 10.3934/medsci.2023007
Fluoxetine is a selective serotonin reuptake inhibitor that is a commonly used drug for the treatment of depression and obsessive-compulsive disorders. Despite the positive effects of this drug, it seems to be associated with various side effects. Genotoxicity or DNA damage is an important side effect of some kinds of drugs. To date, the genotoxicity and cytotoxicity of fluoxetine are partially unknown. In the present study, some oxidative stress methods were used, such as ROS, MDA and GSH evaluation methods in HepG2 cells treated with fluoxetine (1–10 µM). A comet assay was used to evaluate the genotoxic effects of fluoxetine, and flow cytometry was used for apoptosis detection in these hepatic cells. Our data have shown that fluoxetine increased MDA and intracellular concentration of ROS significantly (P < 0.001), while the amount of GSH was reduced significantly (P < 0.001). Our results also indicated that fluoxetine increased the DNA damage of HepG2 cells. The tail percentage of DNA for control cells was 4%, but this percentage was 19%, 28% and 32% for 1, 5 and 10 µM of fluoxetine concentration, respectively (P < 0.01 and P < 0.001). The flow cytometry results have also shown increases in early and late apoptosis for fluoxetine (13.31% and 9.54%, respectively). In conclusion, the present study has shown that fluoxetine is able to induce oxidative stress-dependent DNA damage. Anyway, more studies are needed to accurately explore the molecular and cellular aspects of fluoxetine.

| [1] |
Gheysarzadeh A, Sadeghifard N, Afraidooni L, et al. (2018) Serum-based microRNA biomarkers for major depression: MiR-16, miR-135a, and miR-1202. J Res Med Sci 23: 69. https://doi.org/10.4103/jrms.JRMS_879_17 
|
| [2] |
Alboni S, Van Dijk RM, Poggini S, et al. (2017) Fluoxetine effects on molecular, cellular and behavioral endophenotypes of depression are driven by the living environment. Mol Psychiatry 22: 552-561. https://doi.org/10.1038/mp.2015.142
|
| [3] |
Trivedi MH, Rush AJ, Wisniewski SR, et al. (2006) Evaluation of outcomes with citalopram for depression using measurement-based care in STAR*D: implications for clinical practice. Am J Psychiatry 163: 28-40. https://doi.org/10.1176/appi.ajp.163.1.28
|
| [4] |
Kirsch I, Deacon BJ, Huedo-Medina TB, et al. (2008) Initial severity and antidepressant benefits: a meta-analysis of data submitted to the Food and Drug Administration. PLoS Med 5: e45. https://doi.org/10.1371/journal.pmed.0050045
|
| [5] | Hughes S, Foster RG, Peirson SN, et al. (2016) Inhibitory effects of fluoxetine on photosensitive retinal ganglion cells. Investig Ophthalmol Vis Sci 57: 4660. |
| [6] |
Kaur G, Invally M, Sanzagiri R, et al. (2015) Evaluation of the antidepressant activity of Moringa oleifera alone and in combination with fluoxetine. J Ayurveda Integr Med 6: 273-279. https://doi.org/10.4103/0975-9476.172384
|
| [7] |
Sandanapitchai V, Ramasamy S, Pichai E (2016) Prospective study to evaluate the efficacy of fluoxetine in comparison with amitriptyline in patients with depression. Int J Basic Clin Pharmacol 5: 2552-2555. https://doi.org/10.18203/2319-2003.ijbcp20164122
|
| [8] |
Gheysarzadeh A, Yazdanparast R (2015) STAT5 reactivation by catechin modulates H2O2-induced apoptosis through miR-182/FOXO1 pathway in SK-N-MC cells. Cell Biochem Biophys 71: 649-656. https://doi.org/10.1007/s12013-014-0244-6
|
| [9] |
Mohammadi M, Yazdanparast R (2011) Modulation of H2O2-induced mitogen-activated protein kinases activation and cell death in SK-N-MC cells by EUK134, a salen derivative. Basic Clin Pharmacol Toxicol 108: 378-384. https://doi.org/10.1111/j.1742-7843.2010.00664.x
|
| [10] | Sies H (2000) What is oxidative stress?. Oxidative stress and vascular disease . Boston: Springer 1-8. https://doi.org/10.1007/978-1-4615-4649-8_1 |
| [11] |
Ji C, Rouzer CA, Marnett LJ, et al. (1998) Induction of cell cycle arrest by the endogenous product of lipid peroxidation, malondialdehyde. Carcinogenesis 19: 1275-1283. https://doi.org/10.1093/carcin/19.7.1275
|
| [12] |
Alarifi S, Ali D, Alkahtani S, et al. (2014) Iron oxide nanoparticles induce oxidative stress, DNA damage, and caspase activation in the human breast cancer cell line. Biol Trace Elem Res 159: 416-424. https://doi.org/10.1007/s12011-014-9972-0
|
| [13] |
Gheysarzadeh A, Yazdanparast R (2012) Inhibition of H2O2-induced cell death through FOXO1 modulation by EUK-172 in SK-N-MC cells. Eur J Pharmacol 697: 47-52. https://doi.org/10.1016/j.ejphar.2012.09.036
|
| [14] |
Ghassemi-Barghi N, Varshosaz J, Etebari M, et al. (2016) Role of recombinant human erythropoietin loading chitosan-tripolyphosphate nanoparticles in busulfan-induced genotoxicity: analysis of DNA fragmentation via comet assay in cultured HepG2 cells. Toxicol In Vitro 36: 46-52. https://doi.org/10.1016/j.tiv.2016.07.001
|
| [15] |
Maleki F, Sadeghifard N, Hosseini HM, et al. (2019) Growth-inhibitory effects of TGFαL3-SEB chimeric protein on colon cancer cell line. Biomed Pharmacother 110: 190-196. https://doi.org/10.1016/j.biopha.2018.11.025
|
| [16] |
Zheng Y, Chang X, Huang Y, et al. (2023) The application of antidepressant drugs in cancer treatment. Biomed Pharmacother 157: 113985. https://doi.org/10.1016/j.biopha.2022.113985
|
| [17] |
Szegezdi E, Logue SE, Gorman AM, et al. (2006) Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep 7: 880-885. https://doi.org/10.1038/sj.embor.7400779
|
| [18] |
Rasheva VI, Domingos PM (2009) Cellular responses to endoplasmic reticulum stress and apoptosis. Apoptosis 14: 996-1007. https://doi.org/10.1007/s10495-009-0341-y
|
| [19] |
Carson DA, Ribeiro JM (1993) Apoptosis and disease. Lancet 341: 1251-1254. https://doi.org/10.1016/0140-6736(93)91154-E
|
| [20] |
Birben E, Sahiner UM, Sackesen C, et al. (2012) Oxidative stress and antioxidant defense. World Allergy Organ J 5: 9-19. https://doi.org/10.1097/WOX.0b013e3182439613
|
| [21] |
Laborde E (2010) Glutathione transferases as mediators of signaling pathways involved in cell proliferation and cell death. Cell Death Differ 17: 1373-1380. https://doi.org/10.1038/cdd.2010.80
|
| [22] |
Lima CF, Fernandes-Ferreira M, Pereira-Wilson C (2006) Phenolic compounds protect HepG2 cells from oxidative damage: relevance of glutathione levels. Life Sci 79: 2056-2068. https://doi.org/10.1016/j.lfs.2006.06.042
|
| [23] |
Zlatković J, Todorović N, Tomanović N, et al. (2014) Chronic administration of fluoxetine or clozapine induces oxidative stress in rat liver: a histopathological study. Eur J Pharm Sci 59: 20-30. https://doi.org/10.1016/j.ejps.2014.04.010
|
| [24] |
Kotan VO, Sarandol E, Kirhan E, et al. (2011) Effects of long-term antidepressant treatment on oxidative status in major depressive disorder: a 24-week follow-up study. Prog Neuropsychopharmacol Biol Psychiatry 35: 1284-1290. https://doi.org/10.1016/j.pnpbp.2011.03.021
|
| [25] |
Novío S, Núñez MJ, Amigo G, et al. (2011) Effects of fluoxetine on the oxidative status of peripheral blood leucocytes of restraint-stressed mice. Basic Clin Pharmacol Toxicol 109: 365-371. https://doi.org/10.1111/j.1742-7843.2011.00736.x
|
| [26] |
Zafir A, Banu N (2007) Antioxidant potential of fluoxetine in comparison to Curcuma longa in restraint-stressed rats. Eur J Pharmacol 572: 23-31. https://doi.org/10.1016/j.ejphar.2007.05.062
|
| [27] |
Bowie M, Pilie P, Wulfkuhle J, et al. (2015) Fluoxetine induces cytotoxic endoplasmic reticulum stress and autophagy in triple negative breast cancer. World J Clin Oncol 6: 299-311. https://doi.org/10.5306/wjco.v6.i6.299
|
| [28] |
Ma J, Yang YR, Chen W, et al. (2016) Fluoxetine synergizes with temozolomide to induce the CHOP-dependent endoplasmic reticulum stress-related apoptosis pathway in glioma cells. Oncol Rep 36: 676-684. https://doi.org/10.3892/or.2016.4860
|
| [29] |
Dicks N, Gutierrez K, Michalak M, et al. (2015) Endoplasmic reticulum stress, genome damage, and cancer. Front Oncol 5: 11. https://doi.org/10.3389/fonc.2015.00011
|
| [30] |
Corcoran CA, Luo X, He Q, et al. (2005) Genotoxic and endoplasmic reticulum stresses differentially regulate TRB3 expression. Cancer Biol Ther 4: 1063-1067. https://doi.org/10.4161/cbt.4.10.2205
|




Figures(5)

Somayeh Boshtam, Mohammad Shokrzadeh, Nasrin Ghassemi-Barghi. Fluoxetine induces oxidative stress-dependent DNA damage in human hepatoma cells[J]. AIMS Medical Science, 2023, 10(1): 69-79. doi: 10.3934/medsci.2023007







 DownLoad:
DownLoad: