Citation: Auwal Idris Kabuga, Yusuf Ashiru Hassan, Muhammad Ibrahim Getso. Acute infection with measles virus predisposes to mastoiditis with concomitant facial paralysis and neck abscess: A minireview of pathomechanism and diagnostic approach[J]. AIMS Medical Science, 2020, 7(4): 269-277. doi: 10.3934/medsci.2020016

| [1] |
Sansonetti PJ (2018) Measles 2018: A tale of two anniversaries. EMBO Mol Med 10: e9176. doi: 10.15252/emmm.201809176

|
| [2] |
Onoja A, Adeniji A, Faneye A (2013) Measles complications in a Nigerian hospital setting. Clin Rev Opinions 5: 18-23. doi: 10.5897/CRO12.008
|
| [3] | Duru CO, Peterside O, Adeyemi OO (2014) A 5 year review of childhood measles at the Niger Delta University Teaching Hospital, Bayelsa state, Nigeria. J Med Med Sci 5: 78-86. |
| [4] | Zipprich J, Winter K, Hacker J, et al. (2015) Measles outbreak—California, December 2014–February 2015. MMWR Morb Mortal Wkly Rep 64: 153-154. |
| [5] |
Wendorf KA, Winter K, Zipprich J, et al. (2017) Subacute sclerosing panencephalitis: The devastating measles complication that might be more common than previously estimated. Clin Infect Dis 65: 226-232. doi: 10.1093/cid/cix302
|
| [6] |
Weldegebriel GG, Gasasira A, Harvey P, et al. (2011) Measles resurgence following a nationwide measles vaccination campaign in Nigeria, 2005–2008. J Infect Dis 204: S226-S231. doi: 10.1093/infdis/jir136
|
| [7] |
Singh SK (2014) Human respiratory viral infections CRC Press. doi: 10.1201/b16778
|
| [8] |
Suboti R (1976) Histopathological findings in the inner ear caused by measles. J Laryngol Otol 90: 173-181. doi: 10.1017/S0022215100081937
|
| [9] | Mustafa A, Toçi B, Thaçi H, et al. (2018) Acute mastoiditis complicated with concomitant Bezold's abscess and lateral sinus thrombosis. Case Rep Otolaryngol 2018. |
| [10] |
Leibovitz E (2008) Complicated otitis media and its implications. Vaccine 26: G16-G19. doi: 10.1016/j.vaccine.2008.11.008
|
| [11] |
Tatsuo H, Ono N, Tanaka K, et al. (2000) SLAM (CDw150) is a cellular receptor for measles virus. Nature 406: 893-897. doi: 10.1038/35022579
|
| [12] |
De Witte L, De Vries RD, Van Der Vlist M, et al. (2008) DC-SIGN and CD150 have distinct roles in transmission of measles virus from dendritic cells to T-lymphocytes. PLoS Pathog 4: e1000049. doi: 10.1371/journal.ppat.1000049
|
| [13] |
Buckland R, Wild TF (1997) Is CD46 the cellular receptor for measles virus? Virus Res 48: 1-9. doi: 10.1016/S0168-1702(96)01421-9
|
| [14] |
Noyce RS, Bondre DG, Ha MN, et al. (2011) Tumor cell marker PVRL4 (nectin 4) is an epithelial cell receptor for measles virus. PLoS Pathog 7: e1002240. doi: 10.1371/journal.ppat.1002240
|
| [15] |
Laksono BM, de Vries RD, Duprex WP, et al. (2020) Measles pathogenesis, immune suppression and animal models. Curr Opin Virol 41: 31-37. doi: 10.1016/j.coviro.2020.03.002
|
| [16] |
Laksono BM, Grosserichter-Wagener C, de Vries RD, et al. (2018) In vitro measles virus infection of human lymphocyte subsets demonstrates high susceptibility and permissiveness of both naive and memory B cells. J Virol 92: e00131-18. doi: 10.1128/JVI.00131-18
|
| [17] |
Laksono BM, de Vries RD, Verburgh RJ, et al. (2018) Studies into the mechanism of measles-associated immune suppression during a measles outbreak in the Netherlands. Nat Commun 9: 1-10. doi: 10.1038/s41467-018-07515-0
|
| [18] |
Laksono BM, de Vries RD, McQuaid S, et al. (2016) Measles virus host ınvasion and pathogenesis. Viruses 8: 210. doi: 10.3390/v8080210
|
| [19] |
Perry RT, Halsey NA (2004) The clinical significance of measles: A review. J Infect Dis 189: S4-S16. doi: 10.1086/377712
|
| [20] |
Kurle G (2017) Bezold's abscess: A rare complication of chronic suppurative otitis media in a 13 year old female child. Int J Otorhinolaryngol Head Neck Surg 3: 437. doi: 10.18203/issn.2454-5929.ijohns20171194
|
| [21] |
Oestreicher-Kedem Y, Popovtzer A, Raveh E, et al. (2005) Complications of mastoiditis in children at the onset of a new millennium. Ann Otol Rhinol Laryngol 114: 147-152. doi: 10.1177/000348940511400212
|
| [22] |
Antoli-Candela F, Stewart T (1974) The pathophysiology of otologic facial paralysis. Otolaryngol Clin North Am 7: 309-330. doi: 10.1016/S0030-6665(20)32839-5
|
| [23] | Moreano EH, Paparella MM, Zelterman D, et al. (1994) Prevalence of facial canal dehiscence and of persistent stapedial artery in the human middle ear: A report of 1000 temporal bones. Laryngoscope 104: 309-320. |
| [24] |
Selesnick SH, Jackler RK (1992) Facial paralysis in suppurative ear disease. Oper Tech Otolaryngol Head Neck Surg 3: 61-68. doi: 10.1016/S1043-1810(10)80014-4
|
| [25] |
Elliott CA, Zalzal GH, Gottlieb WR (1996) Acute otitis media and facial paralysis in children. Ann Otol Rhinol Laryngol 105: 58-62. doi: 10.1177/000348949610500110
|
| [26] |
Spiegel JH, Lustig LR, Lee KC, et al. (1998) Contemporary presentation and management of a spectrum of mastoid abscesses. Laryngoscope 108: 822-828. doi: 10.1097/00005537-199806000-00009
|
| [27] |
Marioni G, de Filippis C, Tregnaghi A, et al. (2001) Bezold's abscess in children: Case report and review of the literature. Int J Pediatr Otorhinolaryngol 61: 173-177. doi: 10.1016/S0165-5876(01)00564-X
|
| [28] | Vlastos I, Helmis G, Athanasopoulos I, et al. (2010) Acute mastoiditis complicated with bezold abscess, sigmoid sinus thrombosis and occipital osteomyelitis in a child. Eur Rev Med Pharmacol Sci 14: 635-638. |
| [29] | Nelson D, Jeanmonod R (2013) Bezold abscess: A rare complication of mastoiditis. Am J Emerg Med 31: 1626. e1623-1626. e1624. |
| [30] |
Al-Baharna H, Al-Mubaireek H, Arora V (2016) Bezold's abscess: A case report and review of cases over 14 years. Indian J Otology 22: 148. doi: 10.4103/0971-7749.187978
|
| [31] | Govea-Camacho LH, Pérez-Ramírez R, Cornejo-Suárez A, et al. (2016) Diagnosis and treatment of the complications of otitis media in adults. Case series and literature review. Cirugía Y Cirujanos (English Edition) 84: 398-404. |
| [32] |
McMullan B (2009) Bezold's abscess: A serious complication of otitis media. J Paediatr Child Health 45: 616-618. doi: 10.1111/j.1440-1754.2009.01575.x
|
| [33] | Minor LB, Glasscock ME, Poe D (2010) Glasscock-Shambaugh Surgery of the ear PMPH-USA. |
| [34] |
Yildirim I, Shea KM, Pelton SI (2015) Pneumococcal disease in the era of pneumococcal conjugate vaccine. Infect Dis Clin 29: 679-697. doi: 10.1016/j.idc.2015.07.009
|
| [35] |
Yorgancılar E, Yıldırım M, Gun R, et al. (2013) Complications of chronic suppurative otitis media: A retrospective review. Eur Arch Otorhinolaryngol 270: 69-76. doi: 10.1007/s00405-012-1924-8
|
| [36] |
Secko M, Aherne A (2013) Diagnosis of Bezold abscess using bedside ultrasound. J Emerg Med 44: 670-672. doi: 10.1016/j.jemermed.2012.07.066
|
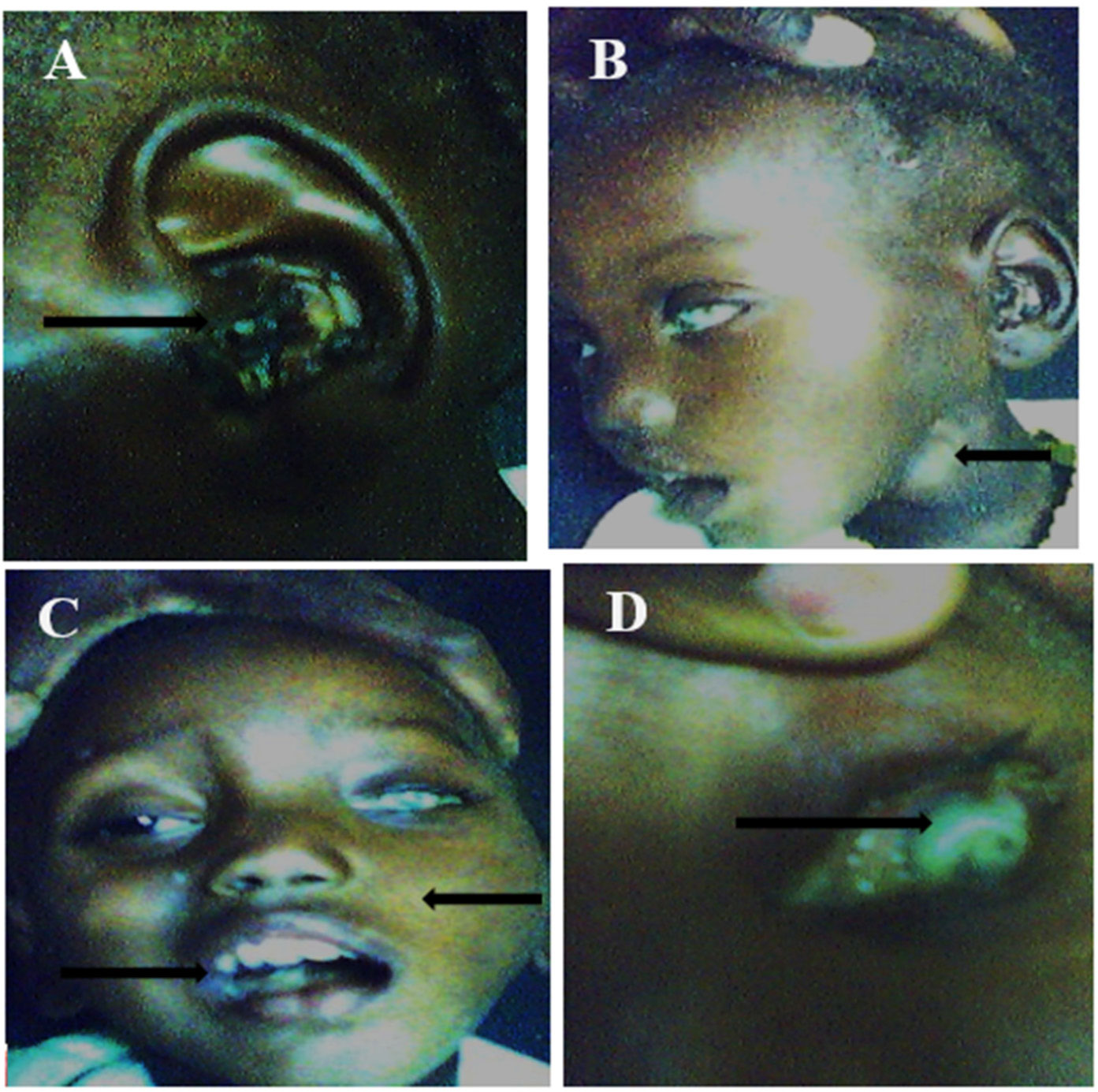

Figures(2)

Auwal Idris Kabuga, Yusuf Ashiru Hassan, Muhammad Ibrahim Getso. Acute infection with measles virus predisposes to mastoiditis with concomitant facial paralysis and neck abscess: A minireview of pathomechanism and diagnostic approach[J]. AIMS Medical Science, 2020, 7(4): 269-277. doi: 10.3934/medsci.2020016







 DownLoad:
DownLoad: