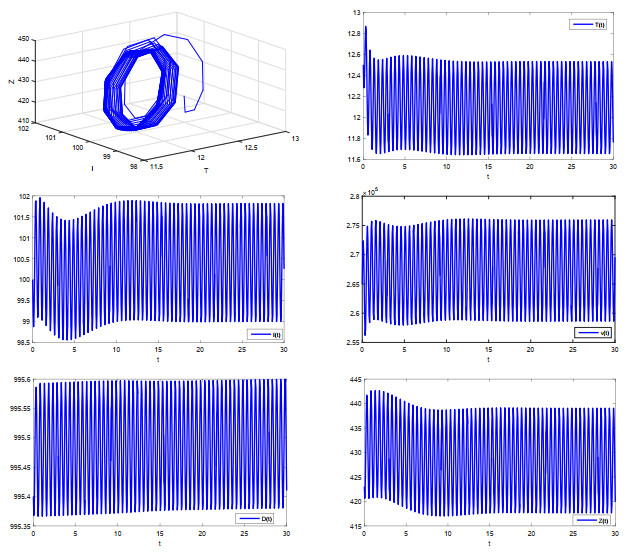
The purpose of this paper is to give some sufficient conditions for the existence of periodic oscillation of a class of in-host MERS-Cov infection model with cytotoxic T lymphocyte (CTL) immune response. A new technique is developed to obtain a lower bound of the state variable characterizing CTL immune response in the model. Our results expand on some previous works.
Citation: Tuersunjiang Keyoumu, Ke Guo, Wanbiao Ma. Periodic oscillation for a class of in-host MERS-CoV infection model with CTL immune response[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 12247-12259. doi: 10.3934/mbe.2022570
The purpose of this paper is to give some sufficient conditions for the existence of periodic oscillation of a class of in-host MERS-Cov infection model with cytotoxic T lymphocyte (CTL) immune response. A new technique is developed to obtain a lower bound of the state variable characterizing CTL immune response in the model. Our results expand on some previous works.

| [1] |
A. M. Zaki, S. van Boheemen, T. M. Bestebroer, A. D. M. E. Osterhauset, R. A. M. Fouchier, Isolation of a novel coronavirus from a man with pneumonia in saudi arabia, N. Engl. J. Med., 367 (2012), 1814–1820. https://doi.org/10.1056/NEJMoa1211721 doi: 10.1056/NEJMoa1211721

|
| [2] |
A. M. Alnuqaydan, A. G. Almutary, A. Sukamaran, B. T. W. Yang, X. T. Lee, W. X. Lim, et al., Middle East Respiratory Syndrome (MERS) virus-pathophysiological axis and the current treatment strategies, AAPS PharmSciTech, 22 (2021), 173. https://doi.org/10.1208/s12249-021-02062-2 doi: 10.1208/s12249-021-02062-2
|
| [3] |
J. Lee, G. Chowell, E. Jung, A dynamic compartmental model for the Middle East respiratory syndrome outbreak in the Republic of Korea: A retrospective analysis on control interventions and superspreading events, J. Theor. Biol., 408 (2016), 118–126. https://doi.org/10.1016/j.jtbi.2016.08.009 doi: 10.1016/j.jtbi.2016.08.009
|
| [4] |
Q. Lin, A. P. Chiu, S. Zhao, D. He, Modeling the spread of Middle East respiratory syndrome coronavirus in Saudi Arabia, Stat. Methods Med. Res., 27 (2018), 1968–1978. https://doi.org/10.1177/0962280217746442 doi: 10.1177/0962280217746442
|
| [5] |
T. Sardar, I. Ghosh, X. Rodó, J. Chattopadhyay, A realistic two-strain model for MERS-CoV infection uncovers the high risk for epidemic propagation, PLoS Negl. Trop. Dis., 14 (2020), e0008065. https://doi.org/10.1371/journal.pntd.0008065 doi: 10.1371/journal.pntd.0008065
|
| [6] |
I. Ghosh, S. S. Nadim, J. Chattopadhyay, Zoonotic MERS-CoV transmission: Modeling, backward bifurcation and optimal control analysis, Nonlinear Dyn., 103 (2021), 2973–2992. https://doi.org/10.1007/s11071-021-06266-w doi: 10.1007/s11071-021-06266-w
|
| [7] |
K. S. Kim, K. Ejima, S. Iwanami, Y. Fujita, H. Ohashi, Y. Koizumi, et al., A quantitative model used to compare within host SARS-CoV-2, MERS-CoV, and SARS-CoV dynamics provides insights into the pathogenesis and treatment of SARS-CoV-2, PLoS Biol., 19 (2021), e3001128. https://doi.org/10.1371/journal.pbio.3001128 doi: 10.1371/journal.pbio.3001128
|
| [8] |
S. Tang, W. Ma, P. Bai, A novel dynamic model describing the spread of the MERS-CoV and the expression of dipeptidyl peptidase 4, Comput. Math. Methods Med., 2017 (2017), 5285810. https://doi.org/10.1155/2017/5285810 doi: 10.1155/2017/5285810
|
| [9] |
M. A. Nowak, C. R. M. Bangham, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74–79. https://doi.org/10.1126/science.272.5258.74 doi: 10.1126/science.272.5258.74
|
| [10] |
A. U. Neumann, N. P. Lam, H. Dahari, D. R. Gretch, T. E. Wiley, T. J. Layden, et al., Hepatitis C viral dynamics in vivo and the antiviral efficacy of interferon-$\alpha$ therapy, Science, 282 (1998), 103–107. https://doi.org/10.1126/science.282.5386.103 doi: 10.1126/science.282.5386.103
|
| [11] |
A. S. Perelson, P. W. Nelson, Mathematical analysis of HIV-1 dynamics in vivo, SIAM Rev., 41 (1999), 3–44. https://doi.org/10.1137/S0036144598335107 doi: 10.1137/S0036144598335107
|
| [12] |
T. Keyoumu, W. Ma, K. Guo, Existence of positive periodic solutions for a class of in-host MERS-CoV infection model with periodic coefficients, AIMS Math., 7 (2021), 3083–3096. https://doi.org/10.3934/math.2022171 doi: 10.3934/math.2022171
|
| [13] | R. E. Gaines, J. L. Mawhin, Coincidence Degree and Nonlinear Differential Equations, Springer-Verlag, Berlin, 1977. https://doi.org/10.1007/BFb0089537 |
| [14] |
C. Egami, Bifurcation analysis of the Nowak-Bangham model in CTL dynamics, Math. Biosci., 221 (2009), 33–42. https://doi.org/10.1016/j.mbs.2009.06.005 doi: 10.1016/j.mbs.2009.06.005
|
| [15] |
J. Pang, J. Cui, J. Hui, The importance of immune responses in a model of hepatitis B virus, Nonlinear Dyn., 67 (2012), 723–734. https://doi.org/10.1007/s11071-011-0022-6 doi: 10.1007/s11071-011-0022-6
|
| [16] |
F. Li, W. Ma, Z. Jiang, D. Li, Stability and Hopf bifurcation in a delayed HIV infection model with general incidence rate and immune impairment, Comput. Math. Method Med., 2015 (2015), 206205. https://doi.org/10.1155/2015/206205 doi: 10.1155/2015/206205
|
| [17] |
J. Li, K. Men, Y. Yang, D. Li, Dynamical analysis on a chronic hepatitis C virus infection model with immune response, J. Theoret. Biol., 365 (2015), 337–346. https://doi.org/10.1016/j.jtbi.2014.10.039 doi: 10.1016/j.jtbi.2014.10.039
|
| [18] |
A. M. Elaiw, N. H. AlShamrani, Stability of HIV/HTLV-I co-infection model with delays, Math Meth Appl Sci., 45 (2022), 238–300. https://doi.org/10.1002/mma.7775 doi: 10.1002/mma.7775
|
| [19] |
G. Li, Y. Fan, Y. Lai, T. Han, Z. Li, P. Zhou, et al., Coronavirus infections and immune responses, J. Med. Virol., 92 (2020), 424–432. https://doi.org/10.1002/jmv.25685 doi: 10.1002/jmv.25685
|
| [20] |
F. Chen, D. Sun, J. Shi, Periodicity in a food-limited population model with toxicants and state dependent delays, J. Math. Anal. Appl., 288 (2003), 136–146. https://doi.org/10.1016/S0022-247X(03)00586-9 doi: 10.1016/S0022-247X(03)00586-9
|
| [21] |
M. Fan, Y. Kuang, Dynamics of a nonautonomous predator-prey system with the Beddington-DeAngelis functional response, J. Math. Anal. Appl., 295 (2004), 15–39. https://doi.org/10.1016/j.jmaa.2004.02.038 doi: 10.1016/j.jmaa.2004.02.038
|
| [22] |
K. Zhao, Y. Li, Four positive periodic solutions to two species parasitical system with harvesting terms, Comput. Math. Appl., 59 (2010), 2703–2710. https://doi.org/10.1016/j.camwa.2010.01.038 doi: 10.1016/j.camwa.2010.01.038
|
| [23] |
A. Muhammadhaji, Z. Teng, Global attractivity of a periodic delayed $N$-species model of facultative mutualism, Discrete Dyn. Nat. Soc., 2013 (2013), 580185. https://doi.org/10.1155/2013/580185 doi: 10.1155/2013/580185
|
| [24] |
C. Celik, N. T. Ozarslan, Existence of periodic solution for a tumor growth model with vaccine interaction, Topol. Methods Nonlinear Anal., 55 (2020), 37–49. https://doi.org/10.12775/TMNA.2019.083 doi: 10.12775/TMNA.2019.083
|
| [25] |
K. Guo, K. Song, W. Ma, Existence of positive periodic solutions of a delayed periodic Microcystins degradation model with nonlinear functional responses, Appl. Math. Lett., 131 (2022), 108056. https://doi.org/10.1016/j.aml.2022.108056 doi: 10.1016/j.aml.2022.108056
|
| [26] |
J. Yang, R. Xu, H. Sun, Dynamics of a seasonal brucellosis disease model with nonlocal transmission and spatial diffusion, Commun. Nonlinear Sci. Numer. Simul., 94 (2021), 105551. https://doi.org/10.1016/j.cnsns.2020.105551 doi: 10.1016/j.cnsns.2020.105551
|
| [27] |
C. Li, J. Xu, J. Liu, Y. Zhou, The within-host viral kinetics of SARS-CoV-2, Math. Biosci. Eng., 17 (2020), 2853–2861. https://doi.org/10.3934/mbe.2020159 doi: 10.3934/mbe.2020159
|
Figures(1)

Tuersunjiang Keyoumu, Ke Guo, Wanbiao Ma. Periodic oscillation for a class of in-host MERS-CoV infection model with CTL immune response[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 12247-12259. doi: 10.3934/mbe.2022570







 DownLoad:
DownLoad: