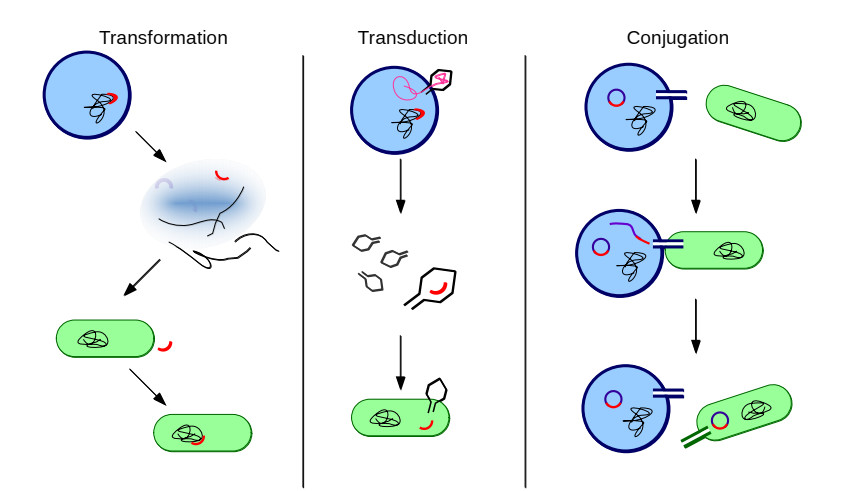
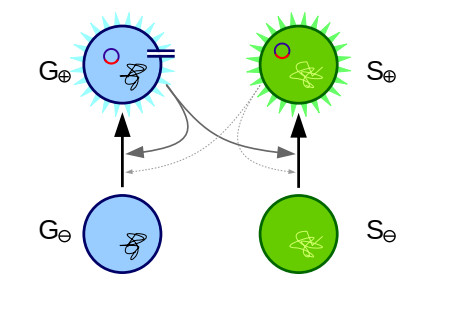
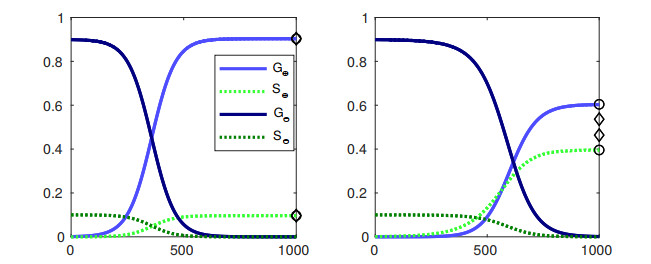
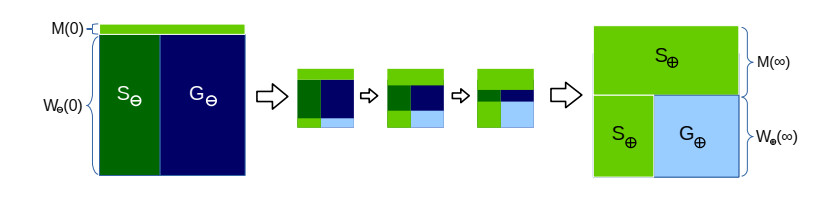
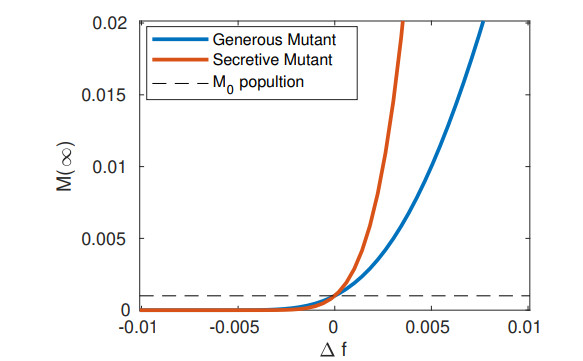
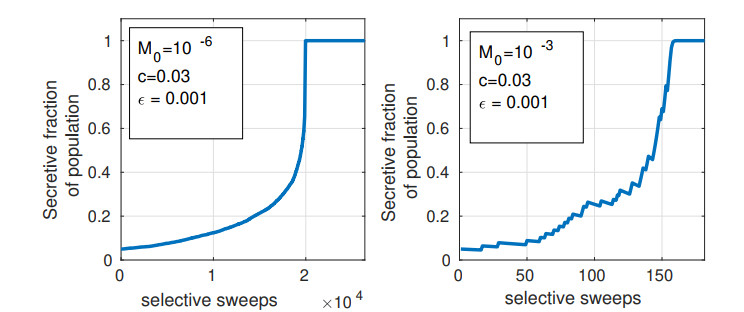
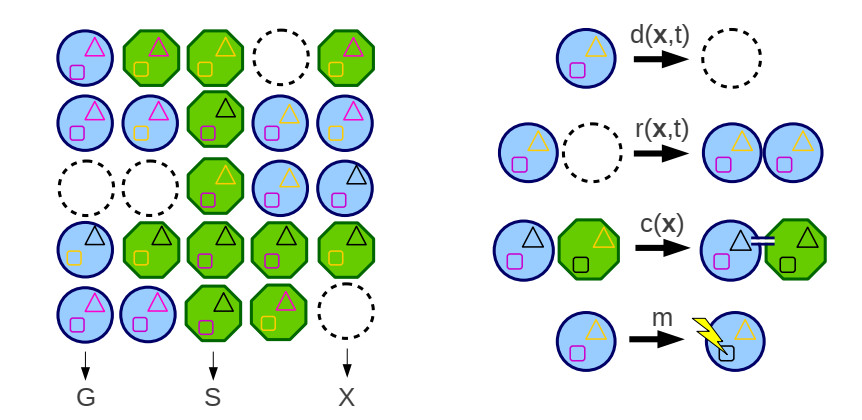
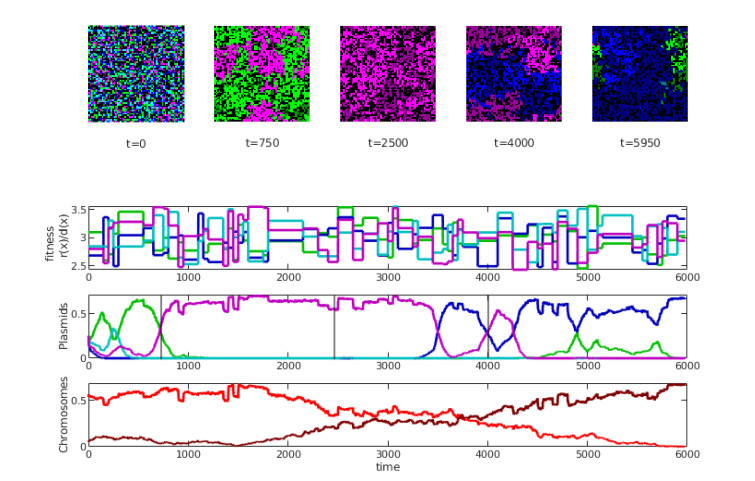
Bacteria, in contrast to eukaryotic cells, contain two types of genes: chromosomal genes that are fixed to the cell, and plasmids, smaller loops of DNA capable of being passed from one cell to another. The sharing of plasmid genes between individual bacteria and between bacterial lineages has contributed vastly to bacterial evolution, allowing specialized traits to 'jump ship' between one lineage or species and the next. The benefits of this generosity from the point of view of both recipient cell and plasmid are generally understood: plasmids receive new hosts and ride out selective sweeps across the population, recipient cells gain new traits (such as antibiotic resistance). Explaining this behavior from the point of view of donor cells is substantially more difficult. Donor cells pay a fitness cost in order to share plasmids, and run the risk of sharing advantageous genes with their competition and rendering their own lineage redundant, while seemingly receiving no benefit in return. Using both compartment based models and agent based simulations we demonstrate that 'secretive' genes which restrict horizontal gene transfer are favored over a wide range of models and parameter values, even when sharing carries no direct cost. 'Generous' chromosomal genes which are more permissive of plasmid transfer are found to have neutral fitness at best, and are generally disfavored by selection. Our findings lead to a peculiar paradox: given the obvious benefits of keeping secrets, why do bacteria share information so freely?
Citation: Alastair D. Jamieson-Lane, Bernd Blasius. The gossip paradox: Why do bacteria share genes?[J]. Mathematical Biosciences and Engineering, 2022, 19(6): 5482-5508. doi: 10.3934/mbe.2022257
Bacteria, in contrast to eukaryotic cells, contain two types of genes: chromosomal genes that are fixed to the cell, and plasmids, smaller loops of DNA capable of being passed from one cell to another. The sharing of plasmid genes between individual bacteria and between bacterial lineages has contributed vastly to bacterial evolution, allowing specialized traits to 'jump ship' between one lineage or species and the next. The benefits of this generosity from the point of view of both recipient cell and plasmid are generally understood: plasmids receive new hosts and ride out selective sweeps across the population, recipient cells gain new traits (such as antibiotic resistance). Explaining this behavior from the point of view of donor cells is substantially more difficult. Donor cells pay a fitness cost in order to share plasmids, and run the risk of sharing advantageous genes with their competition and rendering their own lineage redundant, while seemingly receiving no benefit in return. Using both compartment based models and agent based simulations we demonstrate that 'secretive' genes which restrict horizontal gene transfer are favored over a wide range of models and parameter values, even when sharing carries no direct cost. 'Generous' chromosomal genes which are more permissive of plasmid transfer are found to have neutral fitness at best, and are generally disfavored by selection. Our findings lead to a peculiar paradox: given the obvious benefits of keeping secrets, why do bacteria share information so freely?

| [1] |
A. O. Summers, S. Silver, Mercury Resistance in a Plasmid-Bearing Strain of Escherichia coli, J. Bacteriol., 112 (1972), 1228–1236. https://doi.org/10.1128/jb.112.3.1228-1236.1972 doi: 10.1128/jb.112.3.1228-1236.1972

|
| [2] |
Z. Shao, H. Zhao, H. Zhao, DNA assembler, an in vivo genetic method for rapid construction of biochemical pathways, Nucleic Acids Res., 37 (2009), e16–e16. https://doi.org/10.1093/nar/gkn991 doi: 10.1093/nar/gkn991
|
| [3] | T. J. Johnson, L. K. Nolan, Pathogenomics of the virulence plasmids of escherichia coli, Microbiol. Mol. Biol. Rev., 73 (2009), 750–774. https://dx.doi.org/10.1128%2FMMBR.00015-09 |
| [4] | P. M. Bennett, Plasmid encoded antibiotic resistance: acquisition and transfer of antibiotic resistance genes in bacteria, Br. J. Pharmacol., 153 (2008), S347–S357. https://dx.doi.org/10.1038%2Fsj.bjp.0707607 |
| [5] |
T. Dagan, Y. Artzy-Randrup, W. Martin, Modular networks and cumulative impact of lateral transfer in prokaryote genome evolution, Proc. Natl. Acad. Sci. U.S.A., 105 (2008), 10039–10044. https://doi.org/10.1073/pnas.0800679105 doi: 10.1073/pnas.0800679105
|
| [6] |
X. Yang, E. Wai-Chi Chan, R. Zhang, S. Chen, A conjugative plasmid that augments virulence in klebsiella pneumoniae, Nat. Microbiol., 4 (2019), 2039–2043. https://doi.org/10.1038/s41564-019-0566-7 doi: 10.1038/s41564-019-0566-7
|
| [7] |
K. Chen, E. W. C. Chan, S. Chen, Evolution and transmission of a conjugative plasmid encoding both ciprofloxacin and ceftriaxone resistance in Salmonella, Emerg. Microbes Infect., 8 (2019), 396–403. https://doi.org/10.1080/22221751.2019.1585965 doi: 10.1080/22221751.2019.1585965
|
| [8] | S. Peter, M. Bosio, C. Gross, D. Bezdan, J. Gutierrez, P. Oberhettinger, et. al., Tracking of antibiotic resistance transfer and rapid plasmid evolution in a hospital setting by nanopore sequencing, mSphere, 5. https://doi.org/10.1128/mSphere.00525-20 |
| [9] |
C. M. Johnson, A. D. Grossman, Integrative and conjugative elements (ICEs): What they do and how they work, Annu. Rev. Genet., 49 (2015), 577–601. https://doi.org/10.1146/annurev-genet-112414-055018 doi: 10.1146/annurev-genet-112414-055018
|
| [10] |
W. G. Eberhard, Evolution in bacterial plasmids and levels of selection, Q. Rev. Biol., 65 (1990), 3–22. https://doi.org/10.1086/416582 doi: 10.1086/416582
|
| [11] |
C. M. Thomas, K. M. Nielsen, Mechanisms of, and barriers to, horizontal gene transfer between bacteria, Nat. Rev. Microbiol., 3 (2005), 711–721. https://doi.org/10.1038/nrmicro1234 doi: 10.1038/nrmicro1234
|
| [12] | G. Koraimann, M. A. Wagner, Social behavior and decision making in bacterial conjugation, Front. Cell. Infect. Microbiol., 4. https://dx.doi.org/10.3389%2Ffcimb.2014.00054 |
| [13] |
P. H. Oliveira, M. Touchon, E. P. C. Rocha, Regulation of genetic flux between bacteria by restriction modification systems, Proc. Natl. Acad. Sci. U.S.A., 113 (2016), 5658–5663. https://doi.org/10.1073/pnas.1603257113 doi: 10.1073/pnas.1603257113
|
| [14] |
M. P. Garciláin-Barcia, F. de la Cruz, Why is entry exclusion an essential feature of conjugative plasmids?, Plasmid, 60 (2008), 1–18. https://doi.org/10.1016/j.plasmid.2008.03.002 doi: 10.1016/j.plasmid.2008.03.002
|
| [15] | J. P. J. Hall, A. J. Wood, E. Harrison, M. A. Brockhurst, Source-sink plasmid transfer dynamics maintain gene mobility in soil bacterial communities, Proc. Natl. Acad. Sci. U.S.A., 201600974. https://doi.org/10.1073/pnas.1600974113 |
| [16] |
C. Dahlberg, L. Chao, Amelioration of the cost of conjugative plasmid carriage in Eschericha coli K12, Genetics, 165 (2003), 1641–1649. https://doi.org/10.1093/genetics/165.4.1641 doi: 10.1093/genetics/165.4.1641
|
| [17] |
C. T. Bergstrom, M. Lipsitch, B. R. Levin, Natural selection, infectious transfer and the existence conditions for bacterial plasmids, Genetics, 155 (2000), 1505–1519. https://doi.org/10.1093/genetics/155.4.1505 doi: 10.1093/genetics/155.4.1505
|
| [18] |
M. J. Bottery, A. J. Wood, M. A. Brockhurst, Adaptive modulation of antibiotic resistance through intragenomic coevolution, Nat. Ecol. Evol., 1 (2017), 1364–1369. http://dx.doi.org/10.1038/s41559-017-0242-3 doi: 10.1038/s41559-017-0242-3
|
| [19] |
K. Trautwein, S. E. Will, R. Hulsch, U. Maschmann, K. Wiegmann, M. Hensler, et. al., Native plasmids restrict growth of phaeobacter inhibens DSM 17395: Energetic costs of plasmids assessed by quantitative physiological analyses, Environ. Microbiol., 18 (2016), 4817–4829. https://doi.org/10.1111/1462-2920.13381 doi: 10.1111/1462-2920.13381
|
| [20] | A. San Millan, C. Maclean, Fitness costs of plasmids: A limit to plasmid transmission, Microbiol. Spectr., 5. https://doi.org/10.1128/microbiolspec.mtbp-0016-2017 |
| [21] |
D. A. Baltrus, Exploring the costs of horizontal gene transfer, Trends Ecol., 28 (2013), 489–495. https://doi.org/10.1016/j.tree.2013.04.002 doi: 10.1016/j.tree.2013.04.002
|
| [22] | S. A. Mc Ginty, D. J. Rankin, The evolution of conflict resolution between plasmids and their bacterial hosts, Evolution, 66 (2012), 1662–1670. |
| [23] | J. Colom, D. Batista, A. Baig, Y. Tang, S. Liu, F. Yuan, et. al., Sex pilus specific bacteriophage to drive bacterial population towards antibiotic sensitivity, Sci. Rep., 9 (2019), 1–11. https://www.nature.com/articles/s41598-019-48483-9 |
| [24] |
E. Harrison, M. A. Brockhurst, Plasmid-mediated horizontal gene transfer is a coevolutionary process, Trends Microbiol., 20 (2012), 262–267. https://doi.org/10.1016/j.tim.2012.04.003 doi: 10.1016/j.tim.2012.04.003
|
| [25] |
T. Dimitriu, C. Lotton, J. Bénard-Capelle, D. Misevic, S. P. Brown, A. B. Lindner, et. al., Genetic information transfer promotes cooperation in bacteria, Proc. Natl. Acad. Sci. U.S.A., 111 (2014), 11103–11108. https://doi.org/10.1073/pnas.1406840111 doi: 10.1073/pnas.1406840111
|
| [26] |
T. Dimitriu, D. Misevic, C. Lotton, S. P. Brown, A. B. Lindner, F. Taddei, Indirect fitness benefits enable the spread of host genes promoting costly transfer of beneficial plasmids, PLoS Biol., 14 (2016), e1002478. https://doi.org/10.1371/journal.pbio.1002478 doi: 10.1371/journal.pbio.1002478
|
| [27] |
A. Jamieson-Lane, B. Blasius, Comment on "indirect fitness benefits enable the spread of host genes promoting costly transfer of beneficial plasmids", PLoS Biol., 19 (2021), e3001449. https://doi.org/10.1371/journal.pbio.3001449 doi: 10.1371/journal.pbio.3001449
|
| [28] |
U. Liberman, M. W. Feldman, Modifiers of mutation rate: A general reduction principle, Theor. Popul. Biol., 30 (1986), 125–142. https://doi.org/10.1016/0040-5809(86)90028-6 doi: 10.1016/0040-5809(86)90028-6
|
| [29] | U. Liberman, M. W. Feldman, The reduction principle for genetic modifiers of the migration rate, in Mathematical Evolutionary Theory, Princeton University Press, 2014,111–137. |
| [30] |
U. Liberman, M. W. Feldman, A general reduction principle for genetic modifiers of recombination, Theor. Popul. Biol., 30 (1986), 341–371. https://doi.org/10.1016/0040-5809(86)90040-7 doi: 10.1016/0040-5809(86)90040-7
|
| [31] |
L. Altenberg, M. W. Feldman, Selection, generalized transmission and the evolution of modifier genes. I. the reduction principle, Genetics, 117 (1987), 559–572. https://doi.org/10.1093/genetics/117.3.559 doi: 10.1093/genetics/117.3.559
|
| [32] |
L. Altenberg, U. Liberman, M. W. Feldman, Unified reduction principle for the evolution of mutation, migration, and recombination, Proc. Natl. Acad. Sci. U.S.A., 114 (2017), E2392–E2400. https://doi.org/10.1073/pnas.1619655114 doi: 10.1073/pnas.1619655114
|
| [33] | S. P. Otto, T. Day, A biologist's guide to mathematical modeling in ecology and evolution, Princeton University Press, 2011. |
| [34] | E. L. Ince, Ordinary differential equations, Dover publishing, 1956. |
| [35] | C. M. Thomas, Plasmid Incompatibility, in Molecular Life Sciences: An Encyclopedic Reference(ed. E. Bell), Springer, New York, 2021, 1–3. https://doi.org/10.1007/978-1-4614-6436-5_565-2 |
| [36] |
J. Cullum, P. Broda, Rate of segregation due to plasmid incompatibility, Genet. Res., 33 (1979), 61–79, https://doi.org/10.1017/S0016672300018176 doi: 10.1017/S0016672300018176
|
| [37] | J. R. Scott, Regulation of plasmid replication, Microbiol. Rev., 48 (1984), 1–23. https://dx.doi.org/10.1016%2Fb978-0-12-048850-6.50006-5 |
| [38] | C. Gago-Córdoba, J. Val-Calvo, A. Miguel-Arribas, E. Serrano, P. K. Singh, D. Abia, et.al., Surface exclusion revisited: function related to differential expression of the surface exclusion system of bacillus subtilis plasmid pLS20, Front. Microbiol., 10. https: //doi.org/10.3389/fmicb.2019.01502 |
| [39] |
D. T. Gillespie, Exact stochastic simulation of coupled chemical reactions, J. Phys. Chem., 81 (1977), 2340–2361. https://doi.org/10.1021/j100540a008 doi: 10.1021/j100540a008
|
| [40] | A. Jamieson-Lane, alastair-JL/HGTparadox, 2020, https://github.com/alastair-JL/HGTparadox |
| [41] |
P. a. P. Moran, Random processes in genetics, Math. Proc. Camb. Philos. Soc., 54 (1958), 60–71. https://doi.org/10.1017/S0305004100033193 doi: 10.1017/S0305004100033193
|
| [42] |
E. Lieberman, C. Hauert, M. A. Nowak, Evolutionary dynamics on graphs, Nature, 433 (2005), 312–316. https://doi.org/10.1038/nature03204 doi: 10.1038/nature03204
|
| [43] |
R. Hermsen, J. B. Deris, T. Hwa, On the rapidity of antibiotic resistance evolution facilitated by a concentration gradient, Proc. Natl. Acad. Sci. U.S.A., 109 (2012), 10775–10780. https://doi.org/10.1073/pnas.1117716109 doi: 10.1073/pnas.1117716109
|
| [44] | J. Maynard Smith, What use is sex?, J. Theor. Biol., 30 (1971), 319–335. https://doi.org/10.1016/0022-5193(71)90058-0 |
| [45] | G. C. Williams, Sex and evolution, (MPB-8), Volume 8, Princeton University Press, 2020. |
| [46] | R. Williams, Introduction, in Politics and Technology(ed. R. Williams), Studies in Comparative Politics, Macmillan Education UK, London, 1971, 7–10. https://doi.org/10.1007/978-1-349-01385-2_1 |
| [47] | S. F. Institute, C. G. Langton, Artificial life, volume {I}: Proceedings of an interdisciplinary workshop on synthesis and simulation of living systems, Westview Press, Redwood City, Calif, 1989. |
| [48] |
P. J. Gerrish, R. E. Lenski, The fate of competing beneficial mutations in an asexual population, Genetica, 102 (1998), 127. http://dx.doi.org/10.1023/A:1017067816551 doi: 10.1023/A:1017067816551
|
| [49] |
S. P. Otto, N. H. Barton, The evolution of recombination: Removing the limits to natural selection, Genetics, 147 (1997), 879–906. https://doi.org/10.1093/genetics/147.2.879 doi: 10.1093/genetics/147.2.879
|
| [50] |
N. Colegrave, Sex releases the speed limit on evolution, Nature, 420 (2002), 664–666. https://doi.org/10.1038/nature01191 doi: 10.1038/nature01191
|
| [51] |
E. A. Ostrowski, Enforcing cooperation in the social amoebae, Curr. Biol., 29 (2019), R474–R484. http://dx.doi.org/10.1016/j.cub.2019.04.022 doi: 10.1016/j.cub.2019.04.022
|
| [52] | G. Cordoni, E. Palagi, Back to the future: A glance over wolf social behavior to understand dog-human relationship, Animals, 9. https://doi.org/10.3390/ani9110991 |
| [53] |
T. Verma, N. a. M. Araújo and H. J. Herrmann, Revealing the structure of the world airline network, Sci. Rep., 4 (2014), 5638. http://dx.doi.org/10.1038/srep05638 doi: 10.1038/srep05638
|
| [54] | J. Bell, The interstellar age: Inside the forty-year voyager mission, First edition edition, Dutton, New York, New York, 2015. |
| [55] |
M. A. Nowak, Five rules for the evolution of cooperation, Science, 314 (2006), 1560–1563. https://doi.org/10.1126/science.1133755 doi: 10.1126/science.1133755
|
| [56] |
L. Lehmann, L. Keller, S. West, D. Roze, Group selection and kin selection: Two concepts but one process, Proc. Natl. Acad. Sci. U.S.A., 104 (2007), 6736–6739. https://doi.org/10.1073/pnas.0700662104 doi: 10.1073/pnas.0700662104
|
| [57] |
J. Birch, S. Okasha, Kin selection and its critics, BioScience, 65 (2015), 22–32. https://doi.org/10.1093/biosci/biu196 doi: 10.1093/biosci/biu196
|
| [58] | J. B. S. Haldane, The causes of evolution, London: Longmans, Green, 1932. Available from: http://archive.org/details/causesofevolutio00hald_0 |
| [59] |
T. Dimitriu, D. Misevic, A. B. Lindner, F. Taddei, S. P. Brown, Bacteria can be selected to help beneficial plasmids spread, PLOS Biol., 19 (2021), e3001489. https://doi.org/10.1371/journal.pbio.3001489 doi: 10.1371/journal.pbio.3001489
|
| [60] |
T. Stalder, L. M. Rogers, C. Renfrow, H. Yano, Z. Smith, E. M. Top, Emerging patterns of plasmid-host coevolution that stabilize antibiotic resistance, Sci. Rep., 7 (2017), 4853. https://doi.org/10.1038/s41598-017-04662-0 doi: 10.1038/s41598-017-04662-0
|
| [61] |
T. Kiers, M. Duhamel, Y. Beesetty, J. Mensah, O. Franken, E. Verbruggen, et. al., Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis, Science, 333 (2011), 880–882. https://doi.org/10.1126/science.1208473 doi: 10.1126/science.1208473
|
| [62] |
S. Hortal, K. L. Plett, J. M. Plett, T. Cresswell, M. Johansen, E. Pendall, et. al., Role of plant-fungal nutrient trading and host control in determining the competitive success of ectomycorrhizal fungi, ISME J., 11 (2017), 2666–2676. https://doi.org/10.1038/ismej.2017.116 doi: 10.1038/ismej.2017.116
|
| [63] | M. A. Nowak, K. Sigmund, Evolution of indirect reciprocity, Nature, 437 (2005), 1291–1298. https://doi.org/gdfdgrf |
| [64] |
H. Ohtsuki, Y. Iwasa, How should we define goodness?–reputation dynamics in indirect reciprocity, J. Theor. Biol., 231 (2004), 107–120. https://doi.org/10.1016/j.jtbi.2004.06.005 doi: 10.1016/j.jtbi.2004.06.005
|
| [65] | D. Sun, Pull in and push out: Mechanisms of horizontal gene transfer in bacteria, Front. Microbiol., 9 (2018). https://doi.org/10.3389/fmicb.2018.02154 |
| [66] | M. J. Wade, A critical review of the models of group selection, Q Rev Biol, 53 (1978), 101–114. https://www.journals.uchicago.edu/doi/abs/10.1086/410450 |
| [67] |
A. Traulsen, M. A. Nowak, Evolution of cooperation by multilevel selection, Proc. Natl. Acad. Sci. U.S.A., 103 (2006), 10952–10955. https://doi.org/10.1073/pnas.0602530103 doi: 10.1073/pnas.0602530103
|
| [68] |
L. Altenberg, An evolutionary reduction principle for mutation rates at multiple loci, Bull. Math. Biol., 73 (2011), 1227–1270. https://doi.org/10.1007/s11538-010-9557-9 doi: 10.1007/s11538-010-9557-9
|
| [69] |
F. M. Stewart, B. R. Levin, The population biology of bacterial plasmids: A priori conditions for the existence of conjugationally transmitted factors, Genetics, 87 (1977), 209–228. https://doi.org/10.1093/genetics/87.2.209 doi: 10.1093/genetics/87.2.209
|
| [70] |
S. J. Tazzyman, S. Bonhoeffer, Fixation probability of mobile genetic elements such as plasmids, Theor. Popul. Biol., 90 (2013), 49–55. https://doi.org/10.1016/j.tpb.2013.09.012 doi: 10.1016/j.tpb.2013.09.012
|
| [71] |
L. N. Lili, N. F. Britton, E. J. Feil, The persistence of parasitic plasmids, Genetics, 177 (2007), 399–405. https://doi.org/10.1534/genetics.107.077420 doi: 10.1534/genetics.107.077420
|
| [72] | A. D. Halleran, E. Flores-Bautista, R. M. Murray, Quantitative characterization of random partitioning in the evolution of plasmid-encoded traits, 2019, (Unpublished work) http://resolver.caltech.edu/CaltechAUTHORS:20190402-080939441 |
| [73] |
N. v. d. Hoeven, A mathematical model for the co-existence of incompatible, conjugative plasmids in individual bacteria of a bacterial population, J. Theor. Biol., 110 (1984), 411–423. https://doi.org/10.1016/s0022-5193(84)80183-6 doi: 10.1016/s0022-5193(84)80183-6
|
| [74] |
A. Ilangovan, S. Connery, G. Waksman, Structural biology of the gram-negative bacterial conjugation systems, Trends Microbiol., 23 (2015), 301–310. https://doi.org/10.1016/j.tim.2015.02.012 doi: 10.1016/j.tim.2015.02.012
|
| [75] |
W. Lee, M. van Baalen, V. A. A. Jansen, Siderophore production and the evolution of investment in a public good: an adaptive dynamics approach to kin selection, J. Theor. Biol., 388 (2016), 61–71. https://doi.org/10.1016/j.jtbi.2015.09.038 doi: 10.1016/j.jtbi.2015.09.038
|
| [76] |
J. Smith, The social evolution of bacterial pathogenesis, Proc. Royal Soc. B, 268 (2001), 61–69. https://doi.org/10.1098/rspb.2000.1330 doi: 10.1098/rspb.2000.1330
|
| [77] |
T. Nogueira, D. J. Rankin, M. Touchon, F. Taddei, S. P. Brown, E. P. C. Rocha, Horizontal gene transfer of the secretome drives the evolution of bacterial cooperation and virulence, Curr. Biol., 19 (2009), 1683–1691. https://doi.org/10.1016/j.cub.2009.08.056 doi: 10.1016/j.cub.2009.08.056
|
Figures(9) / Tables(1)

Alastair D. Jamieson-Lane, Bernd Blasius. The gossip paradox: Why do bacteria share genes?[J]. Mathematical Biosciences and Engineering, 2022, 19(6): 5482-5508. doi: 10.3934/mbe.2022257







 DownLoad:
DownLoad: