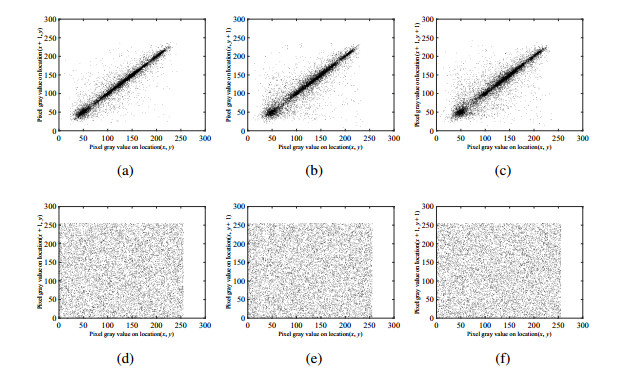
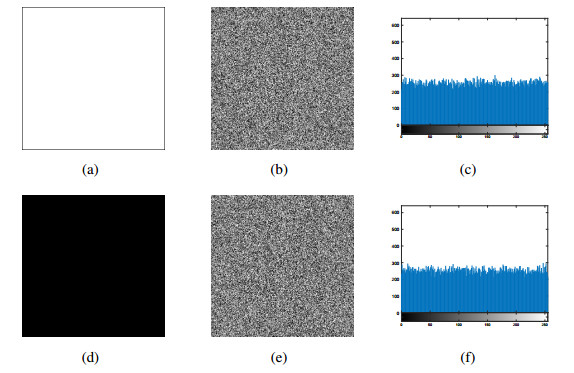
In this paper, an asymmetric image encryption algorithm based on DNA coding and hyperchaotic system is designed. Unlike other image encryption schemes, for example, sharing of same private keys between sender and receiver, and fixed rules with simple operation, three stages are studied as follows to deal with these problems. Firstly, to eliminate the possible risk of key transmission and management, the initial values of the hyperchaotic system are generated for ahead by the RSA (Rivest-Shamir-Adleman) algorithm and the plain image, in which the sum of odd rows, even rows, odd columns, and even columns are computed respectively to extra the plain message from the plain image as input of RSA algorithm. Then, a mathematical map is established to transform all of them into initial values of the hyperchaotic system. Secondly, the pixel level permutation is performed to confuse the image according to the chaotic sequences generated. Finally, to solve the problem of fixed rules with simple operations in current DNA based image encryption algorithms, dynamical DNA encryption is designed to diffuse the permuted image. The process of DNA encryption includes DNA coding, DNA operation and DNA decoding. In particular, DNA rules are selected according to chaotic sequences dynamically, rather than fixed rules with simple operation. Theoretical analysis and numerical simulations show that the proposed algorithm is secure and reliable for image encryption.
Citation: Min Liu, Guodong Ye. A new DNA coding and hyperchaotic system based asymmetric image encryption algorithm[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 3887-3906. doi: 10.3934/mbe.2021194
In this paper, an asymmetric image encryption algorithm based on DNA coding and hyperchaotic system is designed. Unlike other image encryption schemes, for example, sharing of same private keys between sender and receiver, and fixed rules with simple operation, three stages are studied as follows to deal with these problems. Firstly, to eliminate the possible risk of key transmission and management, the initial values of the hyperchaotic system are generated for ahead by the RSA (Rivest-Shamir-Adleman) algorithm and the plain image, in which the sum of odd rows, even rows, odd columns, and even columns are computed respectively to extra the plain message from the plain image as input of RSA algorithm. Then, a mathematical map is established to transform all of them into initial values of the hyperchaotic system. Secondly, the pixel level permutation is performed to confuse the image according to the chaotic sequences generated. Finally, to solve the problem of fixed rules with simple operations in current DNA based image encryption algorithms, dynamical DNA encryption is designed to diffuse the permuted image. The process of DNA encryption includes DNA coding, DNA operation and DNA decoding. In particular, DNA rules are selected according to chaotic sequences dynamically, rather than fixed rules with simple operation. Theoretical analysis and numerical simulations show that the proposed algorithm is secure and reliable for image encryption.

| [1] |
Y. Zhang, Test and verification of AES used for image encryption, 3D Res., 9 (2018), 3. doi: 10.1007/s13319-017-0154-7

|
| [2] |
G. D. Ye, C. Pan, Y. X. Dong, Y. Shi, X. L. Huang, Image encryption and hiding algorithm based on compressive sensing and random numbers insertion, Signal Process, 172 (2020), 107563. doi: 10.1016/j.sigpro.2020.107563
|
| [3] |
L. H. Gong, K. D. Qiu, C. Z. Deng, N. R. Zhou, An image compression and encryption algorithm based on chaotic system and compressive sensing, Opt. Laser Technol., 115 (2019), 257–267. doi: 10.1016/j.optlastec.2019.01.039
|
| [4] | G. D. Ye, C. Pan, Y. X. Dong, K. X. Jiao, X. L. Huang, A novel multi-image visually meaningful encryption algorithm based on compressive sensing and Schur decomposition, Transact. Emerg. Telecommun. Technol., 32 (2021), e4071. |
| [5] |
Z. J. Liu, Q. Guo, L. Xu, M. A. Ahmad, S. T. Liu, Double image encryption by using iterative random binary encoding in gyrator domains, Opt. Express, 18 (2010), 12033–12043. doi: 10.1364/OE.18.012033
|
| [6] |
H. J. Liu, X. Y. Wang, Color image encryption based on one-time keys and robust chaotic maps, Comput. Math. Appl., 59 (2010), 3320–3327. doi: 10.1016/j.camwa.2010.03.017
|
| [7] |
X. Y. Wang, L. T. Liu, Y. Q. Zhang, A novel chaotic block image encryption algorithm based on dynamic random growth technique, Opt. Laser. Eng., 66 (2015), 10–18. doi: 10.1016/j.optlaseng.2014.08.005
|
| [8] | Y. L. Ma, C. Q. Li, B. Ou, Cryptanalysis of an image block encryption algorithm based on chaotic maps, J. Inf. Secur. Appl., 54 (2020), 102566. |
| [9] |
L. F. Liu, S. D. Hao, J. Lin, Z. Wang, X. Y. Hu, S. X. Miao, Image block encryption algorithm based on chaotic maps, IET Signal Process, 12 (2018), 22–30. doi: 10.1049/iet-spr.2016.0584
|
| [10] |
W. H. Liu, K. H. Sun, C. X. Zhu, A fast image encryption algorithm based on chaotic map, Opt. Laser. Eng., 84 (2016), 26–36. doi: 10.1016/j.optlaseng.2016.03.019
|
| [11] |
Z. Y. Hua, Y. C. Zhou, H. J. Huang, Cosine-transform-based chaotic system for image encryption, Inform. Sci., 480 (2019), 403–419. doi: 10.1016/j.ins.2018.12.048
|
| [12] |
M. Alawida, J. S. Teh, A. Samsudin, W. H. Alshoura, An image encryption scheme based on hybridizing digital chaos and finite state machine, Signal Process, 164 (2019), 249–266. doi: 10.1016/j.sigpro.2019.06.013
|
| [13] |
H. J. Li, Y. R. Wang, Z. W. Zuo, Chaos-based image encryption algorithm with orbit perturbation and dynamic state variable selection mechanisms, Opt. Laser. Eng., 115 (2019), 197–207. doi: 10.1016/j.optlaseng.2018.12.002
|
| [14] |
A. Belazi, A. A. A. El-Latif, A. V. Diaconu, R. Rhouma, S. Belghith, Chaos-based partial image encryption scheme based on linear fractional and lifting wavelet transforms, Opt. Laser. Eng., 88 (2017), 37–50. doi: 10.1016/j.optlaseng.2016.07.010
|
| [15] |
M. Kumari, S. Gupta, A. Malik, A superlative image encryption technique based on bit plane using key-based electronic code book, Multimed. Tools Appl., 79 (2020), 33161–33191. doi: 10.1007/s11042-018-6780-0
|
| [16] |
K. Shahna, A. Mohamed, A novel image encryption scheme using both pixel level and bit level permutation with chaotic map, Appl. Soft Comput., 90 (2020), 106162. doi: 10.1016/j.asoc.2020.106162
|
| [17] |
L. Teng, X. Y. Wang, A bit-level image encryption algorithm based on spatio temporal chaotic system and self-adaptive, Opt. Commun., 285 (2012), 4048–4054. doi: 10.1016/j.optcom.2012.06.004
|
| [18] |
X.Y. Wang, H. L. Zhang, A color image encryption with heterogeneous bit-permutation and correlated chaos, Opt. Commun., 342 (2015), 51–60. doi: 10.1016/j.optcom.2014.12.043
|
| [19] |
L. Xu, Z. Li, J. Li, W. Hua, A novel bit-level image encryption algorithm based on chaotic maps, Opt. Laser. Eng., 78 (2016), 17–25. doi: 10.1016/j.optlaseng.2015.09.007
|
| [20] |
Z. L. Zhu, W. Zhang, K. W. Wong, H. Yu, A chaos-based symmetric image encryption scheme using a bit-level permutation, Inform. Sci., 181 (2011), 1171–1186. doi: 10.1016/j.ins.2010.11.009
|
| [21] |
X. L. Chai, Y. R. Chen, L. Broyde, A novel chaos-based image encryption algorithm using DNA sequence operations, Opt. Laser. Eng., 88 (2017), 197–213. doi: 10.1016/j.optlaseng.2016.08.009
|
| [22] |
H. J. Liu, X. Y. Wang, A. kadir, Image encryption using DNA complementary rule and chaotic maps, Appl. Soft Comput., 12 (2012), 1457–1466. doi: 10.1016/j.asoc.2012.01.016
|
| [23] |
D. Ravichandran, A. S. Banu, B. K. Murthy, V. Balasubramanian, S. Fathima, R. Amirtharajan, An efficient medical image encryption using hybrid DNA computing and chaos in transform domain, Med. Biol. Eng. Comput., 59 (2021), 589–605. doi: 10.1007/s11517-021-02328-8
|
| [24] |
X. P. Wei, L. Guo, Q. Zhang, J. X. Zhang, S. G. Lian, A novel color image encryption algorithm based on DNA sequence operation and hyper-chaotic system, J. Syst. Software, 85 (2012), 290–299. doi: 10.1016/j.jss.2011.08.017
|
| [25] |
R. Enayatifar, F. G. Guimaraes, P. Siarry, Index-based permutation-diffusion in multiple-image encryption using DNA sequence, Opt. Laser. Eng., 115 (2019), 131–140. doi: 10.1016/j.optlaseng.2018.11.017
|
| [26] |
M. Yildirim, DNA encoding for RGB image encryption with memristor based neuron model and chaos phenomenon, Microelectron. J., 104 (2020), 104878. doi: 10.1016/j.mejo.2020.104878
|
| [27] |
Q. Zhang, L. Guo, X. P. Wei, Image encryption using DNA addition combining with chaotic maps, Math. Comput. Model., 52 (2010), 2028–2035. doi: 10.1016/j.mcm.2010.06.005
|
| [28] |
H. Hermassi, A. Belazi, R. Rhouma, S. M. Belghith, Security analysis of an image encryption algorithm based on a DNA addition combining with chaotic maps, Multimed. Tools Appl., 72 (2014), 2211–2224. doi: 10.1007/s11042-013-1533-6
|
| [29] |
S. Q. Zhu, C. X. Zhu, W. H. Wang, A new image encryption algorithm based on chaos and secure hash sha-256, Entropy, 20 (2018), 716. doi: 10.3390/e20090716
|
| [30] |
E. E. García-Guerrero, E. Inzunza-González, O. R. López-Bonilla, J. R. Cárdenas-Valdez, E. Tlelo-Cuautle, Randomness improvement of chaotic maps for image encryption in a wireless communication scheme using PIC-microcontroller via Zigbee channels, Chaos Soliton. Fract., 133 (2020), 109646. doi: 10.1016/j.chaos.2020.109646
|
| [31] |
E. Rodríguez-Orozco, E. E. García-Guerrero, E. Inzunza-Gonzalez, O. R. López-Bonilla, A. Flores-Vergara, J. R. Cárdenas-Valdez, et al., FPGA-based chaotic cryptosystem by using voice recognition as access key, Electronics, 7 (2018), 414. doi: 10.3390/electronics7120414
|
| [32] |
C. Tanougast, Hardware implementation of chaos based cipher: Design of embedded systems for security applications, Chaos-Based Cryptography, 354 (2011), 297–330. doi: 10.1007/978-3-642-20542-2_9
|
| [33] |
A. Flores-Vergara, E. E. García-Guerrero, E. Inzunza González, O. R. López-Bonilla, E. Rodrguez-Orozco, J. R. Cárdenas-Valdez, et al., Implementing a chaotic cryptosystem in a 64-bit embedded system by using multiple-precision arithmetic, Nonlinear Dynam., 96 (2019), 497-516. doi: 10.1007/s11071-019-04802-3
|
| [34] |
S. F. Yousif, A. J. Abboud, H. Y. Radhi, Robust image encryption with scanning technology, the El-Gamal algorithm and chaos theory, IEEE Access, 8 (2020), 155184–155209. doi: 10.1109/ACCESS.2020.3019216
|
| [35] | X. Z. Dong, L. Zhang, X. W. Gao, An efficient FPGA implementation of ECC modular inversion over F256, Proceedings of the 2nd International Conference on Cryptography, Security and Privacy, (2018), 29–33. |
| [36] | X. M. Huang, W. Wang, A novel and efficient design for an RSA cryptosystem with a very large key size, IEEE T. Circuits-II, 62 (2015), 972–976. |
| [37] |
S. D. Thabah, M. Sonowal, R. U. Ahmed, P. Saha, Fast and area efficient implementation of RSA algorithm, Proceed. Computer Sci., 165 (2019), 525–531. doi: 10.1016/j.procs.2020.01.024
|
| [38] | G. C. Marchesan, N. R. Weirich, E. C. Culau, I. I. Weber, F. G. Moraes, E. Carara, et al., Exploring RSA performance up to 4096-bit for fast security processing on a flexible instruction set architecture processor, IEEE International Conference on Electronics, Circuits and Systems (ICECS), (2018), 18420517. |
| [39] | T. Adiono, H. Ega, H. Kasan, S. Fuada, S. Harimurti, Full custom design of adaptable montgomery modular multiplier for asymmetric RSA cryptosystem, International Symposium on Intelligent Signal Processing and Communication Systems (ISPACS), (2017), 17524004. |
| [40] |
H. Chen, C. Tanougast, Z. J. Liu, B. Y. Hao, Securing color image by using hyperchaotic system in gyrator transform domains, Opt. Quant. Electron., 48(2016), 396. doi: 10.1007/s11082-016-0669-9
|
| [41] |
N. Zhu, Y. T. Wang, J. Liu, J. H. Xie, H. Zhang, Optical image encryption based on interference of polarized light, Opt. Express, 17 (2009), 13418–13424. doi: 10.1364/OE.17.013418
|
| [42] |
G. D. Ye, K. X. Jiao, H. S. Wu, C. Pan, X. L. Huang, An asymmetric image encryption algorithm based on a fractional-order chaotic system and the RSA public-key cryptosystem, Int. J. Bifurcat. Chaos, 30 (2020), 2050233. doi: 10.1142/S0218127420502338
|
| [43] | J. S. Teh, M. Alawida, Y. C. Sii, Implementation and practical problems of chaos-based cryptography revisited, J. Inf. Secur. Appl., 50 (2020), 102421. |
| [44] |
T. Wang, M. H. Wang, Hyperchaotic image encryption algorithm based on bit-level permutation and DNA encoding, Opt. Laser Technol., 132 (2020), 106355. doi: 10.1016/j.optlastec.2020.106355
|
| [45] | R. S. Ye, Y. T. Xi, Y. L. Ma, A chaotic image encryption scheme using swapping based confusion approach, IEEE international conference on computer communication and the internet, (2016), 16525393. |
| [46] |
X. L. Chai, Z. H. Gan, M. H. Zhang, A fast chaos-based image encryption scheme with a novel plain image-related swapping block permutation and block diffusion, Multimed. Tools Appl., 76 (2017), 15561–15585. doi: 10.1007/s11042-016-3858-4
|
| [47] |
X. L. Xue, D. S. Zhou, C. J. Zhou, New insights into the existing image encryption algorithms based on DNA coding, PLoS One, 15 (2020), e0241184. doi: 10.1371/journal.pone.0241184
|


Figures(8) / Tables(12)

Min Liu, Guodong Ye. A new DNA coding and hyperchaotic system based asymmetric image encryption algorithm[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 3887-3906. doi: 10.3934/mbe.2021194







 DownLoad:
DownLoad: