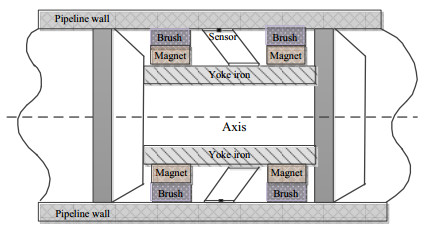
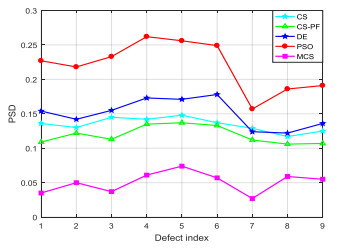
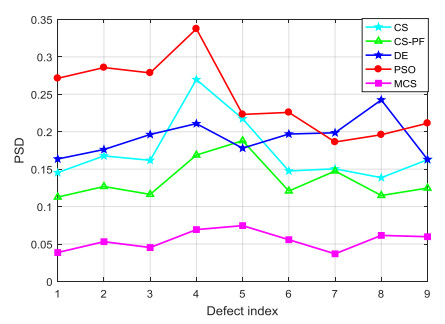
Accurate and efficient estimation for defect profile of magnetic flux leakage (MFL) signals is important for nondestructive evaluation in industry. To improve the accuracy of defect profile reconstruction, an improved reconstruction method based on modified cuckoo search (CS), called MCS, is proposed in this paper. Firstly, a novel single-dimension updating evolution strategy is proposed to avoid the interference between multiple dimensions, which can make full use of the appropriate nest position in the historical search. Secondly, an adaptive multi-strategy difference evolution is introduced into the evolution process to improve the diversity and efficiency of CS algorithm. The proportion factor of each strategy in multi-strategy difference evolution is adjusted dynamically according to the value of the objective fitness. Finally, various MFL signals are selected to verify the effectiveness of the proposed MCS algorithm. The experiment results illustrate that the proposed method has high performance on the quality of the solution and robustness for noise.
Citation: Daqian Zhang, Chen Huang, Jiyou Fei. Defect reconstruction from magnetic flux leakage measurements employing modified cuckoo search algorithm[J]. Mathematical Biosciences and Engineering, 2021, 18(2): 1898-1925. doi: 10.3934/mbe.2021099
Accurate and efficient estimation for defect profile of magnetic flux leakage (MFL) signals is important for nondestructive evaluation in industry. To improve the accuracy of defect profile reconstruction, an improved reconstruction method based on modified cuckoo search (CS), called MCS, is proposed in this paper. Firstly, a novel single-dimension updating evolution strategy is proposed to avoid the interference between multiple dimensions, which can make full use of the appropriate nest position in the historical search. Secondly, an adaptive multi-strategy difference evolution is introduced into the evolution process to improve the diversity and efficiency of CS algorithm. The proportion factor of each strategy in multi-strategy difference evolution is adjusted dynamically according to the value of the objective fitness. Finally, various MFL signals are selected to verify the effectiveness of the proposed MCS algorithm. The experiment results illustrate that the proposed method has high performance on the quality of the solution and robustness for noise.

| [1] | J. Wu, Y. Sun, Y. Kang, Y. Yang, Theoretical analyses of MFL signal affected by discontinuity orientation and sensor-scanning direction, IEEE Trans. Magn., 51 (2015), 1–7. |
| [2] |
J. Pavo, Numerical calculation method for pulsed eddy-current testing, IEEE Trans. Mag., 38 (2002), 1169–1172. doi: 10.1109/20.996299

|
| [3] | B. Hartmut, P. Konstantin, M. Judith, Lorentz force eddy current testing: A novel NDE-technique, Compel Int. J. Comput. Math. Electri. Electron. Eng., 33 (2013), 1965–1977. |
| [4] | Y. Shi, C. Zhang, R. Li, M. Cai, G. Jia, Theory and application of magnetic flux leakage pipeline detection, Sensors, 15 (2015), 31036–31055 |
| [5] |
E. G. Bazulin, A.V. Goncharsky, S. Y. Romanov, Inverse problems of ultrasonic tomography in nondestructive testing: mathematical methods and experiment, Russ. J. Nondestr. Test., 55 (2019), 453–462. doi: 10.1134/S1061830919060020
|
| [6] | W. Han, J. Xu, M. Zhou, M. Zhou, G. Tian, P. Wang, et al., Cuckoo search and particle filter-based inversing approach to estimating defects via magnetic flux leakage signals, IEEE Trans. Magn., 52 (2016), 1–11. |
| [7] | Y. Li, G. Y. Tian, S. Ward, Numerical simulation on magnetic flux leakage evaluation at high speed, NDT & E Int., 39 (2006), 367–373. |
| [8] |
J. Chen, S. Huang, W. Zhao, Equivalent MFL model of pipelines for 3-D defect reconstruction using simulated annealing inversion procedure, Int. J. Appl. Electromagn. Mech., 47 (2015), 551–561. doi: 10.3233/JAE-140022
|
| [9] |
S. M. Dutta, F. H. Ghorbel, R. K. Stanley, Dipole modeling of magnetic flux leakage, IEEE Trans. Magn., 45 (2009), 1959–1965. doi: 10.1109/TMAG.2008.2011895
|
| [10] |
P. Ramuhalli, L. Udpa, Electromagnetic NDE signal inversion by function-approximation neural network, IEEE Trans. Magn., 38 (2002), 3633–3642. doi: 10.1109/TMAG.2002.804817
|
| [11] |
R. H. Priewald, C. Magele, P. D. Ledger, N. R. Pearson, J. S. D. Mason, Fast magnetic flux leakage signal inversion for the reconstruction of arbitrary defect profiles in steel using finite elements, IEEE Trans. Magn., 49 (2013), 506–516. doi: 10.1109/TMAG.2012.2208119
|
| [12] | X. Song, S. Huang, W. Zhao, Quantitative interpretation for the magnetic flux leakage testing data based on neural network, J. Wuhan Univ. Technol., 28 (2006), 443–447. |
| [13] |
K. C. Hari, M. Nabi, S. V. Kulkarni, Improved FEM model for defect-shape construction from MFL signal by using genetic algorithm, IET Sci. Meas. Technol., 1 (2007), 196–200. doi: 10.1049/iet-smt:20060069
|
| [14] |
E. Kurtulus, A. R. Yıldız, S. M. Sait, S. Bureerat, A novel hybrid Harris hawks- simulated annealing algorithm and RBF-based metamodel for design optimization of highway guardrails, Materials Test., 62 (2020), 251–260. doi: 10.3139/120.111478
|
| [15] |
F. M. Li, J. Feng, H. Zhang, J. Liu, S. Lu, D. Ma, Quick reconstruction of arbitrary pipeline defect profiles from MFL measurements employing modified harmony search algorithm, IEEE Trans. Instrum. Meas., 67 (2018), 2200–2213. doi: 10.1109/TIM.2018.2813839
|
| [16] | Y. Zhang, Z. Ye, C. Wang, A fast method for rectangular crack sizes reconstruction in magnetic flux leakage testing, NDT & E Int., 42 (2009), 369–375. |
| [17] | D. Dhiman, K. K. Singh, A. Slowik, V. Chang, A. R. Yildiz, A. Kaur, et al., EMoSOA: a new evolutionary multi-objective seagull optimization algorithm for global optimization, Int. J. Mach. Learn. Cybern., 12 (2021), 571–596. |
| [18] | P. Champasak, N. Panagant, N. Pholdee, S. Bureerat, A. R. Yildiz, Self-adaptive many-objective meta-heuristic based on decomposition for many-objective conceptual design of a fixed wing unmanned aerial vehicle, Aerosp. Sci. Technol., 100 (2020), 105783. |
| [19] |
B. S. Yıldız, A. R. Yıldız, E. I. Albak, H. Abderazek, S. M. Sait, S. Bureerat, Butterfly optimization algorithm for optimum shape design of automobile suspension components, Materials Test., 62 (2020), 365–370. doi: 10.1515/mt-2020-620406
|
| [20] | W. Deng, J. Xu, H. Zhao, Y. Song, A novel gate resource allocation method using improved PSO-based QEA, IEEE Trans. Intel. Transp. Syst., 2020. |
| [21] | Y. Song, D. Wu, W. Deng, X. Gao, T. Li, B. Zhang, et al, MPPCEDE: Multi-population parallel co-evolutionary differential evolution for parameter optimization, Energy Convers. Manage., 228 (2021), 113661. |
| [22] | W. Deng, J. Xu, Y. Song, H. Zhao, An effective improved co-evolution ant colony optimization algorithm with multi-strategies and its application, Int. J. Bio-Inspired Comput., 16 (2020): 158–170. |
| [23] |
M. A. E. Aziz, A. E. Hassanien, Modified cuckoo search algorithm with rough sets for feature selection, Neural Comput. Appl., 29 (2018), 925–934. doi: 10.1007/s00521-016-2473-7
|
| [24] |
Z. Cao, C. Lin, M. Zhou, R. Huang, Scheduling semiconductor testing facility by using Cuckoo search algorithm with reinforcement learning and surrogate modeling, IEEE Trans. Autom. Eng., 16 (2019), 825–837. doi: 10.1109/TASE.2018.2862380
|
| [25] |
I. Durgun, A. R. Yildiz, Structural design optimization of vehicle components using cuckoo search algorithm, Materialprufung/Materials Test., 54 (2012), 185–188. doi: 10.3139/120.110317
|
| [26] | J. Ding, Q. Wang, Q. Zhang, Q. Ye, Y. Ma, A hybrid particle swarm optimization-cuckoo search algorithm and its engineering applications, Math. Probl. Eng., 2019 (2019), 5213759. |
| [27] | A. M. Kamoona, J. C. Patra, A novel enhanced cuckoo search algorithm for contrast enhancement of gray scale images, Appl. Soft Comput., 85 (2019), 105749. |
| [28] | W. Deng, J. J. Xu, Y. J. Song, H. M. Zhao, Differential evolution algorithm with wavelet basis function and optimal mutation strategy for complex optimization problem, Appl. Soft Comput., 100 (2020), 106724. |
| [29] |
J. Zhang, A. C. Sanderson, JADE: Adaptive differential evolution with optional external archive, IEEE Trans. Evol. Comput., 13 (2009), 945–958. doi: 10.1109/TEVC.2009.2014613
|
| [30] | T. Back., Evolutionary Algorithms in Theory and Practice: Evolution Strategies, Evolutionary Programming, Genetic Algorithms, Oxford University Press, Oxford, 1996. |























Figures(26) / Tables(7)

Daqian Zhang, Chen Huang, Jiyou Fei. Defect reconstruction from magnetic flux leakage measurements employing modified cuckoo search algorithm[J]. Mathematical Biosciences and Engineering, 2021, 18(2): 1898-1925. doi: 10.3934/mbe.2021099







 DownLoad:
DownLoad: