Citation: Fahad Al Basir, Yasuhiro Takeuchi, Santanu Ray. Dynamics of a delayed plant disease model with Beddington-DeAngelis disease transmission[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 583-599. doi: 10.3934/mbe.2021032

| [1] | J. D. G. Jones, J. L. Dangl, The plant immune system, Nature, 444 (2006), 323–329. |
| [2] | R. C. Gergerich, V. V. Dolja, Introduction to plant viruses, the invisible foe, Plant Health Instr., 4 (2006). |
| [3] |
G. Neofytou, Y. N. Kyrychko, K.B. Blyuss, Time-delayed model of immune response in plants, J. Theor. Biol., 389 (2016), 28–39. doi: 10.1016/j.jtbi.2015.10.020

|
| [4] | M. Leclerc, T. Doré, C. A. Gilligan, P. Lucas, J. A. N. Filipe, Estimating the delay between host infection and disease (incubation period) and assessing its significance to the epidemiology of plant diseases, PLOS ONE, 9 (2014), 1–15. |
| [5] | M. J. Jeger, L. V. Madden, F. van den Bosch, Plant Virus Epidemiology: applications and prospects for mathematical modeling and analysis to improve understanding and disease control, Plant Dis., 102 (2018), 837–854. |
| [6] | R. P. Almeida, Ecology of emerging vector-borne plant diseases, in Institute of medicine forum on vector-borne diseases: understanding the environmental, human health, and ecological connections., National Academies Press, (2008), 70–77. |
| [7] |
M. J. Jeger, J. Holt, F. van Den Bosch, L.V. Madden, Epidemiology of insect-transmitted plant viruses: modelling disease dynamics and control interventions, Physiol. Entomol., 29 (2004), 291– 304. doi: 10.1111/j.0307-6962.2004.00394.x
|
| [8] |
J. R. Beddington, Mutual interference between parasites or predators and its effect on searching efficiency, J. Anim. Ecol., 44 (1975), 331–340. doi: 10.2307/3866
|
| [9] | D. L. DeAngelis, R. A. Goldstein, R.V. O'Neill, A model for trophic interaction, Ecology, 56 (1975), 881–892. |
| [10] |
X. Song, A. Neumann, Global stability and periodic solution of the viral dynamics, J. Math. Anal. Appl., 329 (2007), 281–297. doi: 10.1016/j.jmaa.2006.06.064
|
| [11] | R. Xu. Global stability of an HIV-1 infection model with saturation infection and intracellular delay, J. Math. Anal. Appl., 375 (2011), 75–81. |
| [12] | G. Huang, W. Ma, Y. Takeuchi, Global analysis for delay virus dynamics model with Beddington– DeAngelis functional response, Appl. Math. Lett., 24 (2011) 1199–1203. |
| [13] |
G. Huang, W. Ma, Y. Takeuchi, Global properties for virus dynamics model with Beddington– DeAngelis functional response, Appl. Math. Lett., 22 (2009), 1690–1693. doi: 10.1016/j.aml.2009.06.004
|
| [14] |
J. Holt, M. J. Jeger, J. M. Thresh, G. W. Otim-Nape, An epidemiological model incorporating vector population dynamics applied to African cassava mosaic virus disease, J. Appl. Ecol., 34 (1997), 793–806. doi: 10.2307/2404924
|
| [15] |
E. Venturino, P. K. Roy, F. A. Basir, A. Datta, A model for the control of the mosaic virus disease in jatropha curcas plantations, Energy Ecol. Environ., 1 (2016), 360–369. doi: 10.1007/s40974-016-0033-8
|
| [16] | B. Buonomo, M. Cerasuolo, Stability and bifurcation in plant-pathogens interactions, Appl. Math. Comput., 232 (2014), 858–871. |
| [17] |
M. Jackson, B. M. Chen-Charpentier, Modeling plant virus propagation with delays, J. Comput. Appl. Math., 309 (2017), 611–621. doi: 10.1016/j.cam.2016.06.009
|
| [18] |
M. Jackson, B. M. Chen-Charpentier, A model of biological control of plant virus propagation with delays, J. Comput. Appl. Math., 330 (2018), 855–865. doi: 10.1016/j.cam.2017.01.005
|
| [19] | J. E. van der Plank, Plant Diseases: Epidemics and Control, Academic Press, 1963. |
| [20] | T. Zhang, X. Meng, Y. Song, Z. Li, Dynamical analysis of delayed plant disease models with continuous or impulsive cultural control strategies, Abstr. Appl. Anal., 2012 (2012). |
| [21] | X. Meng, Z. Li, The dynamics of plant disease models with continuous and impulsive cultural control strategies, J. Theor. Biol., 266 (2010), 2940. |
| [22] | M. Jackson, B. M. Chen-Charpentier, Modeling plant virus propagation with delays, J. Comput. Appl. Math, , 309 (2016), 611–621. |
| [23] | R. Shi, H. Zhao, S. Tang, Global dynamic analysis of a vector-borne plant disease model, Adv. Differ. Equations., 59 (2014). |
| [24] |
M. Banerjee, Y. Takeuchi, Maturation delay for the predators can enhance stable coexistence for a class of prey-predator models, J. Theor. Biol., 412 (2017), 154–171. doi: 10.1016/j.jtbi.2016.10.016
|
| [25] |
B. Buonomo, M. Cerasuolo, The effect of time delay in plant-pathogen interactions with host demography, Math. Biosci. Eng., 12 (2015), 473–490. doi: 10.3934/mbe.2015.12.473
|
| [26] |
F. A. Basir, S. Ray, Impact of incubation delay in plant-vector interaction, Math. Comput. Simul., 170 (2020), 16–31. doi: 10.1016/j.matcom.2019.09.001
|
| [27] |
N. Rakshit, F. A. Basir, A. Banerjee, S. Ray, Dynamics of plant mosaic disease propagation and the usefulness of roguing as an alternative biological control, Ecol. Complex., 38 (2019), 15–23. doi: 10.1016/j.ecocom.2019.01.001
|
| [28] |
F. A. Basir, K. B. Blyuss, S. Ray, Modelling the effects of awareness-based interventions to control the mosaic disease of Jatropha curcas, Ecol. Complex., 36 (2018), 92–100. doi: 10.1016/j.ecocom.2018.07.004
|
| [29] |
J. M. Heffernan, R. J. Smith, L. M. Wahl, Perspectives on the basic reproductive ratio, J. R. Soc. Interface, 2 (2005), 281–93. doi: 10.1098/rsif.2005.0042
|
| [30] |
E. X. DeJesus, C. Kaufman, Routh-Hurwitz criterion in the examination of eigenvalues of a system of nonlinear ordinary differential equations, Phys. Rev. A., 35 (1987), 5288–5290. doi: 10.1103/PhysRevA.35.5288
|
| [31] |
D. W. M. Cook, P. J. Long, S. Ganesh, The combined effect of delayed application of yeast biocontrol agents and fruit curing for the inhibition of the postharvest pathogen Botrytis cinerea in kiwifruit, Postharvest Biol. Technol., 16 (1999), 233–243. doi: 10.1016/S0925-5214(99)00003-4
|
| [32] | D. Liu, K. G. Raghothama, P. M. Hasegawa, R. A. Bressan, Osmotin overexpression in potato delays development of disease symptoms, Proceedings of the National Academy of Sciences of the United States of America, 2012. |
| [33] | K. Niehaus, D. Kapp, A. Pühler, Plant defence and delayed infection of alfalfa pseudonodules induced by an exopolysaccharide (EPS I)-deficient Rhizobium meliloti mutant, Planta, 190 (1993) 415–425. |
| [34] | K. B. Blyuss, F. Al Basir, V. A. Tsygankova, L. O. Biliavska, G. O. Iutynska, S. N. Kyrychko, et al., Control of mosaic disease using microbial biostimulants: insights from mathematical modelling, Ric. Mat., 69 (2020), 437–455. |
| [35] | F. A. Basir, S. Adhurya, M. Banerjee, E. Venturino, S. Ray, modelling the effect of incubation and latent periods on the dynamics of vector-borne plant viral diseases, Bull. Math. Biol., 82 (2020), 94. |
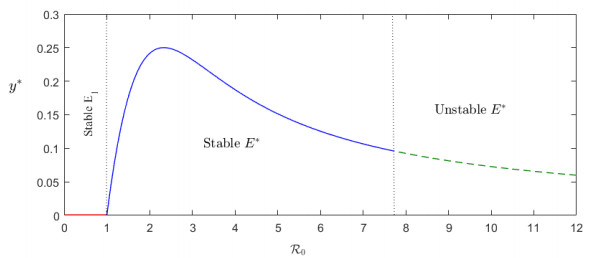
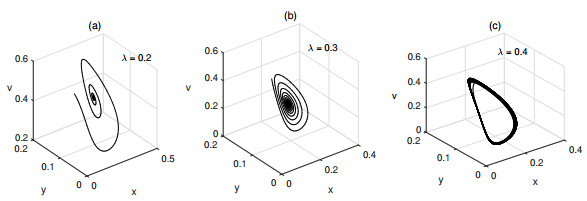
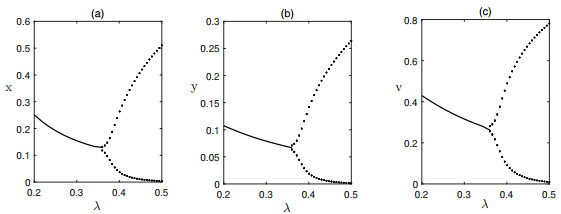
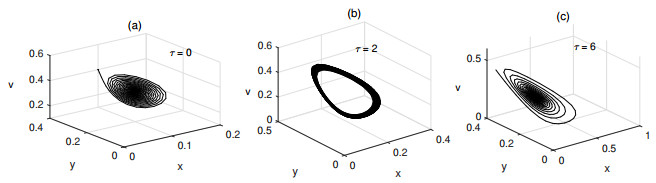
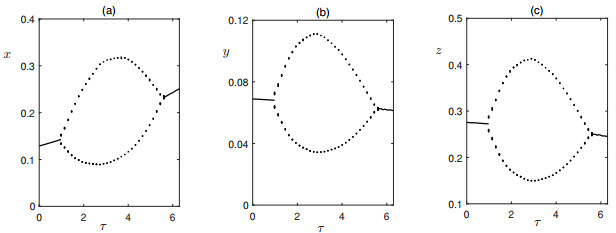
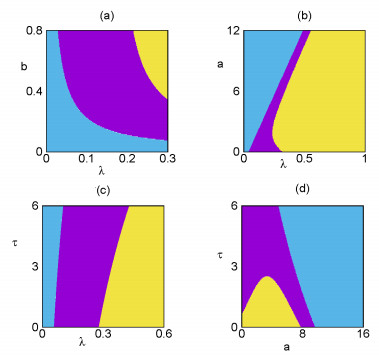
Figures(6) / Tables(1)

Fahad Al Basir, Yasuhiro Takeuchi, Santanu Ray. Dynamics of a delayed plant disease model with Beddington-DeAngelis disease transmission[J]. Mathematical Biosciences and Engineering, 2021, 18(1): 583-599. doi: 10.3934/mbe.2021032







 DownLoad:
DownLoad: