Citation: Yuhua Long, Yining Chen. Global stability of a pseudorabies virus model with vertical transmission[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5234-5249. doi: 10.3934/mbe.2020283

| [1] | N. Ketusing, A. Reeves, K. Portacci, T. Yano, F. Olea-Popelka, T Keefe, et al., Evaluation of strategies for the eradication of pseudorabies virus (Aujeszky's disease) in commercial swine farms in Chiang-Mai and Lampoon Provinces, Thailand, Using a Simulation Disease Spread Model, Transbound. Emerg. Dis., 61(2014), 169-176. |
| [2] | G. Wittmann, H. Rziha, Aujeszky's Disease (Pseudorabies) in Pigs, Springer, Boston, MA, 1989. |
| [3] | J. Zimmerman, L. Karriker, A. Ramirez, K. Schwartz, G. Stevenson, Diseases of Swine 10th Edition, Wiley, 2019. |
| [4] | S. Brogger, Systems analysis in tuberculosis control: A model, Am. Rev. Respir. Dis., 95(1967), 419-434. |
| [5] | S. Z. Jin, L. J. Sun, Q. P. Tong, Z. W. Tong, Clinical diagnosis and ccontrol of Swine Pseudorabies (in Chinese), Shanghai Animal Husbandry and Veterinary Communication, 000(005)(2013), 90- 91. |
| [6] | A. C. Aujeszky, Ueber eine new Infektions krankheit bei Haustieren, Zentbl Bakt ParasitKde, 32(1902), 353-357. |
| [7] | R. P. Hanson, The history of pseudorabies in the United States, J. Am. Vet. Med. Assoc., 124(1954), 259-261. |
| [8] | T. Mettenleiter, Aujeszky, Disease (Pseudorabies) virus: The Virus and molecular pathogenesisState of the Art, June 1999, Vet. Res., 31(2000), 99. |
| [9] | T. Q. An, J. M. Peng, Z. J. Tian, H. Y. Zhao, N. Li, Y. M. Liu, et al., Pseudorabies virus variant in Bartha-K61-vaccinated pigs, China, 2012, Emerg. Infect. Dis., 19(2013), 1749-1755. |
| [10] | N. Denzin, F. J. Conraths, T. C. Mettenleiter, C. M. Freuling, M. Thomas, Monitoring of pseudorabies in wild boar of Germany: A spatiotemporal snalysis, Pathogens, 9(2020). |
| [11] | X. F. Zhai, W. Zhao, K. M. Li, C. Zhang, Genome characteristics and evolution of pseudorabies virus strains in eastern China from 2017 to 2019, Virol. Sinica, 34(2019), 601-609. |
| [12] | H. J. Wu, D. C. Ye, Epidemiological characteristics, purification techniques and application demonstration of pseudorabies in pigs, China Animal Husbandry and Veterinary Digest, 34(2018), 148-148. |
| [13] | C. Freuling, T. Müller, T. Mettenleiter, Vaccines against pseudorabies virus (PrV), Vet. Microbiol., 206(2017), 3-9. |
| [14] | Y. H. Long, L. Wang, Global Dynamics of a delayed Two-patch discrete SIR disease model, Commun. Nonlinear Sci. Numer. Simul., 83(2020), 105-117. |
| [15] | C. H. Li, Y. Wang, F. Y. Jiang, J. H. Hu, Research progress of swine pseudorabies (in Chinese), Progress Animal Med., 29(2008), 68-72. |
| [16] | F. Rao, P. Mandal, Y. Kang, Complicated endemics of an SIRS model with a generalized incidence under preventive vaccination and treatment controls, Appl. Math. Model., 67(2019), 38-61. |
| [17] | O. Diekmann, J. A. P. Heesterbeek, J. A. J. Metz, On the definition and the computation of the basic reproduction ratio ℜ<0 in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28(1990), 365-382. |
| [18] | P. Van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180(2002), 29-48. |
| [19] | J. P. LaSalle, The stability of dynamical systems, Regional Conference Series in Applied Mathematics, Siam, Philadelphia, 1976. |
| [20] | J. Li, Z. Ma, F. Zhang, Stability analysis for an epidemic model with stage structure, Nonlin. Anal., 9(2008), 1672-1679. |
| [21] | Z. E. Ma, Y. C. Zhou, Qualitative and stability methods of ordinary differential equations (in Chinese), Science Press, 2015. |
| [22] | M. Y Li, J. S. Muldowney, Global stability for the SEIR model in epidemiology, Math. Biosci., 125(1995), 155-164. |
| [23] | M. Y. Li, H. L. Smith, L. C. Wang, Global dynamics of an SEIR epidemic model with vertical transmission, SIAM J. Appl. Math., 62(2001), 58-69. |
| [24] | M. Y. Li, J. R. Graef, L. C. Wang, J. Karsai, Global dynamics of a SEIR model with varying total population size, Math. Biosci., 160(1999), 191-213. |
| [25] | H. L. Smith, Systems of Ordinary Differential Equations Which Generate an Order Preserving Flow, A Survey of Results, Society for Industrial and Applied Mathematics, 1988. |
| [26] | M. W. Hirsch, Systems of differential equations which are competitive or cooperative: I. limit sets, SIAM J. Math. Anal., 13(1982), 167-179. |
| [27] | J. S. Muldowney, Compound matrices and ordinary differential equations, Rocky Mountain J. Math., 20(1990), 857-872. |
| [28] | Jr. R. H. Martin, Logarithmic norms and projections applied to linear differential systems, J. Appl. Math Anal. Appl, 45(1974), 432-454. |
| [29] | J. S. Muldowney, Dichotomies and asymptotic behaviour for linear differential systems, Trans. Amer. Math. Soc., 283(1984), 465-484. |
| [30] | L. Cesari, Asymptotic behavior and stability problems in ordinary differential equations, Springer Science & Business Media, 2012. |
| [31] | M. Y. Li, J. S. Muldowney, A geometric approach to Global-stability problems, SIAM J. Math. Anal., 27(1996), 1070-1083. |
| [32] | G. Butler, P. Waltman, Persistence in dynamical systems, J. Differ. Equat., 63(1986), 255-263. |
| [33] | H. W. Hethcote, The mathematics of infectious diseases, SIAM Rev., 42(2000), 599-653. |
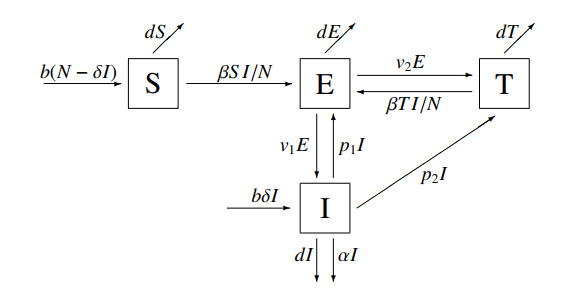
Figures(1)

Yuhua Long, Yining Chen. Global stability of a pseudorabies virus model with vertical transmission[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 5234-5249. doi: 10.3934/mbe.2020283







 DownLoad:
DownLoad: